

 证件证明翻译
证件证明翻译 医疗病历翻译
医疗病历翻译 法律合同翻译
法律合同翻译 公司介绍翻译
公司介绍翻译 学术论文翻译
学术论文翻译 留学移民翻译
留学移民翻译 求职简历翻译
求职简历翻译 产品说明书翻译
产品说明书翻译
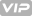




基于11个基因特征指数,可预测结直肠癌的系统性复发(下
3.5. CRC中CMS与PI的比较
我们对PI系统和CMS进行了比较分析,国际协会19对CRC的四个亚类进行了描述:CMS1具有超突变,微卫星不稳定和强免疫激活作用;带有WNT和MYC信号激活的CMS2;CMS3具有明显的代谢失调;以及具有TGF-β激活,基质侵袭和血管生成的CMS4。当在CIT队列中将CMS与PI系统进行比较时,CMS4中的大多数患者(126名中的116名,占92.1%),其中CRC患者具有明显的无复发和总体生存较差的特征,并且往往被诊断为PI将更高级的阶段(III和IV)19归类为高风险(补充图S4a)。在CMS中,CMS1中40.4%(89例中的36例)患者与免疫逃逸的激活和较高的组织病理学分级有关,也被PI归为高危亚组。尽管CMS4和CMS1,CMS2和CMS3高风险CRC患者占主要地位,但与其他亚型相比,低风险CRC的患病率更高:高风险CRC的35.6%(230的82)和24.2%(66的16)分别包括在CMS2和CMS3中(补充图S4a)。当按PI评分对患者进行分类时,得分较高的CRC患者倾向于按CMS4分类(图5)。(图4a),4a),表明由PI分层的高风险亚组可以很好地反映CRC的分子亚型,显示出从上皮-间充质转化(EMT)19带来的不良预后。
的亚型,按预后指数(PI)分层的亚组的比较。.png")
图4根据11个基因签名和4个衍生自共有分子分子亚型(CMS)的亚型,按预后指数(PI)分层的亚组的比较。
PI在(a)CIT,(b)AUS和(c)AMC队列中显示基因表达模式。红色和绿色分别反映了高和低的表达水平
我们还将CMS联盟提供的CMS分类器应用于来自AUS队列的基因表达数据。与CIT队列相似,绝大多数CMS4患者是高危CRC患者(60名中的45名,占75%;补充图S4b)。我们还观察到,CMS1亚组中包含许多高危CRC患者(51个中的23个,占45.1%),而CMS2和CMS3包含的CRC高危患者较少(17.3%(75个中的13个)和7.5%(3) (分别为40)分别与CIT队列中的CMS分类保持一致(补充图S4b)。根据PI评分对患者进行分类时,许多得分较高的CRC患者被归类为CMS4(图2)。(图4b),4b),验证PI分类的预后相关性可作为CRC患者预后不良的指标。
使用构成PI系统的11个基因签名,我们还为AMC队列中的每个患者计算了PI。当PI将AMC队列中的患者分为低风险或高风险亚组时,两种风险亚型之间的疾病复发频率显着不同(Fisher精确检验,P <0.001,优势比= 4.759,95% CI = 1.78–8.64;图图4c),4c),与使用1160个系统性复发相关基因的分层聚类分析相一致(图。 (图。1)。1个)。比较亚组之间的11个基因的表达水平,我们观察到这些基因有显着差异(两次样本t检验,每个P <0.05;补充图S5)。我们还将CMS分类器应用于AMC队列中的RNA-seq数据。检查参与CMS亚组的PI分类高风险CRC患者的频率时,我们获得的比例分别为31%(29个中的9个),44.1%(59个中的26个),50%(18个中的9个)和87.5% CMS1,CMS2,CMS3和CMS4子组中分别包含的百分比(24个中的21个)高风险CRC,与CIT和AUS队列相比,表明CMS4中有类似的高风险患者受累(补充图S4c)。通过PI分类的许多高分率CRC患者显然与CMS4相关,尽管其他CMS亚组中的许多患者被预测为系统性复发的高风险CRC(图。(图4c4c)。
最后,我们比较了PI和CMS分类器之间的基因。基于随机森林预测模型19比较PI系统中的11个基因和CMS分类器中的273个基因时,有趣的是,尽管PI评分高的CRC患者与CMS4的预后相似,但在两个基因列表之间未发现共有基因。亚组。
4. 讨论
大规模高通量技术的广泛进步为CRC提供了许多见解。除了在CRC分子亚型的各种调查,13,23 - 25,由一个国际财团与聚集先前报道亚型努力达成共识也有报道19旨在揭示肿瘤的异质性,并为CRC提供适当的治疗选择。尽管进行了这些严格的努力,但仍很少有可靠的生物标记物可用于准确预测CRC的疾病复发,这是处理或治疗CRC患者的未满足需求之一。在这里,我们收集了总计130例有或没有系统性复发的原发性CRC,并基于RNA-seq分析生成了转录组数据集。通过基因表达谱分析和途径富集分析,我们确定了127个系统性复发相关基因。在这些基因中,有11个基因AK2,BID,CDC25A,EIF4A2,HSPB1,ITGB1,MAP4K4RT-PCR分析证实,MMP12,PTGES3,RHOC和TERF2IP是与多个患者队列中CRC无复发生存相关的最佳特征。PI与这11个基因结合在一起,可强烈预测高风险CRC,并且独立于当前可用的临床指标。生存结果强调了III期CRC患者的预后潜力以及II期CRC患者辅助化疗的特异性。
由于一半的CRC病例与致命事件有关,因此迫切需要对复发或转移进行预测。在这方面,我们认为分子驱动方法将导致易于复发的CRC患者的识别和个性化的治疗选择,通过减少系统性复发来提高生存率。最近,CRC子分类协会(https://github.com/Sage-Bionetworks/crcsc)将六个独立的分类器划分为四个CMS。同样,我们根据当前的PI标准对高危亚组的结果显示,与其他相比,CMS4(间质型)的总体生存率和无复发生存率较差,而CMS1(MSI免疫型)的复发后生存率较差。亚型19。尽管几项随访研究得出了相应的结果,但另一项研究警告说,未考虑肿瘤异质性的各个亚型可能导致将患者分为不适当的亚组26。但是,已经研究了单个或多个标记子集,以阐明CRC中的复发预测因子。MACC1过表达通常导致差的存活结果和被识别为在CRC复发的患者的独立预后指标19,27。
尽管对CRC进行了广泛的研究,但由于CRC的异质性,仍然很难确定将从辅助化疗获益最大或最少的患者。辅助化疗的优势已经确立患者的AJCC III期癌症的21,22,而其在AJCC阶段成效II患者仍存在争议5。在本研究的几个子集分析中,PI系统显示了II期和III期CRC患者的强大预后价值,以及II期CRC患者辅助化疗的显着预测价值。在这项研究中,PI评分可以确定患有II期CRC的低危患者亚组,这些亚组将从辅助化疗中受益。在交互作用分析中,如PI预测,对于低危亚组的患者,化疗与II期密切相关,并且与较差的结果显着相关,而对于高危组的患者,化疗的效果并不明显。 II期疾病。一线化疗显然对PI低表达的II期肿瘤的疗效不及所有其他亚组。因此,其他疗法,包括针对性的生物制剂,
目前提出的PI系统是由11种基因的表达水平组合产生的,这11种基因的表达与CRC的发生和发展密切相关。AK2编码的蛋白质是在线粒体28的膜间空间中发现的一种酶。在人类中AK2缺乏引起的造血缺陷和免疫缺陷29,30。BID,一种BH3相互作用域死亡激动剂,编码一种蛋白质,该蛋白质是细胞死亡调节剂BCL-2家族的成员,其是caspase-8诱导的线粒体损伤的介体。出价在细胞中还具有许多主要活性,例如凋亡,细胞死亡,活化或增殖。虽然之间几个关联BID和癌症已经研究,评价和评估BID在人结肠疾病,如结肠直肠腺瘤或结肠炎,已经证明31,32,这表明BID可以是CRC的一个可能的调节。CDC25A,细胞分裂周期25A,在响应DNA损伤后特别降解,导致细胞周期停滞和成纤维细胞与致癌RAS 33相互作用的转化。EIF4A2编码蛋白,真核翻译起始因子4A II,它是EIF2信号传导途径中蛋白质合成起始过程中eIF4F复合物的组成部分。尽管少数细胞功能EIF4A2已经发现,之间的正关联EIF4A2和各种癌症,包括CRC,已经研究34,35,这表明EIF4A2可以是CRC复发的一种新颖的介体。ITGB1编码的蛋白质(Integrin beta-1)是一种膜受体,参与多种过程中的细胞粘附和识别,包括胚胎发生,止血,组织修复,肿瘤细胞的免疫应答和转移扩散。描述ITGB1与各种癌症(例如胃癌36,乳腺癌37和肺癌38)之间显着正相关的大量研究清楚地支持了ITGB1在癌症中的侵略性特征。的先前报道的功能角色ITGB1在CRC细胞系,如细胞粘附39,40,增殖41,细胞凋亡42,和磷酸化43,显然支持ITGB1对CRC侵略性的贡献。MAP4K4编码蛋白MEK激酶激酶4,它与多种生理过程有关,包括细胞迁移,增殖和粘附。CRC的不良预后和疾病进展与MAP4K4表达水平密切相关44。RHOC编码小GTP酶的Rho家族的成员(即ras同源家族C)。RHOC也参与mTOR信号通路,其中mTORC2通过Gho加载Rho家族45的蛋白质来调节细胞骨架组织。红十字会过表达与肿瘤细胞的增殖和转移有关,特别是与CRC转移信号通路有关,强调了含PI的RHOC表达的预后价值。在签名的11个基因中,MMP12(基质金属蛋白酶12),PTGES3(前列腺素E合酶3)和TERF2IP(端粒重复结合因子2相互作用蛋白1)参与某些临床活动46,但它们的临床关联性与CRC的报道很少,这提示这些基因可能是CRC预后的新标记。
尽管我们在多个患者CRC队列中研究了我们PI系统的临床相关性,但我们的研究存在一些局限性。由于AMC发现队列中数据的随访时间短,因此无法在该队列中确认签名中11个基因的预后相关性。尽管这些结果说明了II期CRC患者的化学特异性,但接受辅助化疗的患者数量不足以清楚地确定PI系统的化学反应。另外,由于在PCR验证中仅评估了独立患者组中的样品太少,因此难以确定基因的重要性。PI系统应在较大的独立临床队列中进行严格验证,对CRC患者的随访时间较长。
总之,我们确定了由CRC中11个基因的表达所定义的不同预后亚组。作为预后和预测指标,新近确定的PI系统不仅可以识别高危II期和III期CRC患者,还可以预测受益于辅助化疗最少的低危II期CRC患者。但是,对于PI的实际临床应用,需要更复杂,更严格的验证步骤,这些步骤不仅会评估其他更大的患者队列,而且还会通过液体活检来评估可检测性。
参考
1. Torre LA等人。全球癌症统计资料,2012年。CACancer J. Clin。2015; 65:87-108。doi:10.3322 / caac.21262。
2. Hari DM等。AJCC癌症分期手册第七版结肠癌标准:复杂的修饰是否改善预后评估?J.上午 Coll。手术 2013; 217:181-190。doi:10.1016 / j.jamcollsurg.2013.04.018。
3. Carlsson U,Lasson A,EkelundG。直肠癌根治性手术后的复发率,特别是其准确性。Dis。结肠直肠。1987年;30:431–434。doi:10.1007 / BF02556491。
4. Midgley R,Kerr D.大肠癌。柳叶刀。1999年;353:391-399。doi:10.1016 / S0140-6736(98)07127-X。
5. Varghese A. II期结肠的化学疗法。癌症临床。结肠直肠手术。2015; 28:256-261。doi:10.1055 / s-0035-1564430。
6. Zarour LR等。大肠癌肝转移:范式的发展和未来的方向。细胞分子 胃肠酸。肝素。2017; 3:163-173。doi:10.1016 / j.jcmgh.2017.01.006。
7. Zabaleta J等。结直肠癌患者肺转移切除术后的生存:切除肝转移的病史是否会使预后恶化?文献综述。癌症生物学。中 2017; 14:281–286。doi:10.20892 / j.issn.2095-3941.2017.0073。
8.法基赫(Fakih MG)。转移性结直肠癌:现状和未来方向。J.Clin。Oncol。2015; 33:1809–1824。doi:10.1200 / JCO.2014.59.7633。
9. Galandiuk S,等人。结肠癌和直肠癌根治性切除后的复发方式。手术 Gynecol。Obstet。1992年;174:27–32。
10. Kim JC等人。GSN和OAS2在结直肠癌转移,介导神经周和淋巴血管浸润中的功能相反。一等奖。2018; 13:e0202856。doi:10.1371 / journal.pone.0202856。
11. Benson AB,3rd等。NCCN准则见解:结肠癌,版本2.2018。J.Natl Compr。Canc。网络 2018; 16:359–369。doi:10.6004 / jnccn.2018.0021。]
12. Kim SK等。基于十九个基因的风险评分分类器可预测结直肠癌患者的预后。大声笑 Oncol。2014; 8:1653-1666。doi:10.1016 / j.molonc.2014.06.016。
13.玛丽莎L,等人。结肠癌的基因表达分类分为分子亚型:表征,验证和预后价值。公共科学图书馆 2013; 10:e1001453。doi:10.1371 / journal.pmed.1001453。
14. Jorissen RN。等人。与转移相关的基因表达变化预示了公爵B期和C期大肠癌患者的预后不良。临床 癌症研究。2009; 15:7642-7651。doi:10.1158 / 1078-0432.CCR-09-1431。
15.白先生S,等人。一种多基因检测方法,可预测他莫昔芬治疗的淋巴结阴性乳腺癌的复发率。N. Engl。J. Med。2004; 351:2817-2826。doi:10.1056 / NEJMoa041588。
16. Kim SM等。基于六十五个基因的风险评分分类器可预测肝细胞癌的总体生存率。肝病学。2012; 55:1443-1452。doi:10.1002 / hep.24813。
17. Venables WN,Ripley BD,Venables WN。 S.第四版的《现代应用统计》。纽约:施普林格;2002年。
18. Kim SK,Hwan Kim J,Yun SJ,Kim WJ,Kim SY。APPEX:用于鉴定癌症预后基因表达特征的分析平台。生物信息学。2014; 30:3284-3286。doi:10.1093 / bioinformatics / btu521。
19. Guinney J,等。大肠癌的共有分子亚型。纳特 中 2015; 21:1350–1356。doi:10.1038 / nm.3967。
20. Babjuk M等人。EAU关于非肌肉浸润性膀胱尿路上皮癌的指南:2013年更新。Eur。乌鲁尔。2013; 64:639-653。doi:10.1016 / j.eururo.2013.06.003。
21.劳瑞JA等人。大肠癌的外科手术辅助治疗:左旋咪唑以及左旋咪唑和氟尿嘧啶的联合评估。北部中部癌症治疗小组和梅奥诊所。J.Clin。Oncol。1989年;7:1447–1456。doi:10.1200 / JCO.1989.7.10.1447。
22. Moertel CG,等人。左旋咪唑和氟尿嘧啶用于切除结肠癌的辅助治疗。N. Engl。J. Med。1990年;322:352–358。doi:10.1056 / NEJM199002083220602。
23.癌症基因组图谱,北。人类结肠癌和直肠癌的全面分子表征。自然。2012; 487:330-337。doi:10.1038 / nature11252。
24. Sadanandam A等。一种将细胞表型和对治疗的反应相关联的大肠癌分类系统。纳特 中 2013; 19:619–625。doi:10.1038 / nm.3175。
25. De Sousa EMF等。预后差的结肠癌由分子上不同的亚型定义,并从锯齿状前体病变发展而来。纳特 中 2013; 19:614–618。doi:10.1038 / nm.3174。
26.邓恩PD,等人。挑战癌症分子分层法则:肿瘤内异质性破坏了大肠癌的共识分子亚型和潜在的诊断价值。临床 癌症研究。2016; 22:4095–4104。doi:10.1158 / 1078-0432.CCR-16-0032。
27. Stein U等人。MACC1是新近确定的HGF-MET信号转导的关键调节因子,可预测结肠癌的转移。纳特 中 2009; 15:59-67。doi:10.1038 / nm.1889。
28. Kohler C等人。凋亡过程中从线粒体膜间空间释放腺苷酸激酶2。FEBS Lett。1999年;447:10-12。doi:10.1016 / S0014-5793(99)00251-3。
29. Pannicke U等。网状细胞发育不全(白细胞增多)是由编码线粒体腺苷酸激酶2的基因突变引起的。基因 2009; 41:101-105。doi:10.1038 / ng.265。
30. Lagresle-Peyrou C等。人腺苷酸激酶2缺乏症会引起与感觉神经性耳聋有关的严重造血缺陷。纳特 基因 2009; 41:106-111。doi:10.1038 / ng.278。
31. Heijink DM,等人。一种生物信息学和功能方法,用于识别大肠癌化学预防的新策略。癌基因。2011; 30:2026-2036。doi:10.1038 / onc.2010.578。
32. Yeretssian G等。BID在炎症和先天免疫中的非凋亡作用。自然。2011; 474:96-99。doi:10.1038 / nature09982。
33. Kerkhoff E,Rapp UR。Ras / Raf信号传导的细胞周期目标。癌基因。1998年;17:1457–1462。doi:10.1038 / sj.onc.1202185。
34. Kandoth C等人。跨12种主要癌症类型的突变情况和意义。自然。2013; 502:333-339。doi:10.1038 / nature12634。
35. Saaf AM等。体外极化Caco-2肠上皮细胞的全球转录程序与正常结肠癌和结肠癌中的基因表达程序之间存在平行性。大声笑 生物学 细胞。2007年;18:4245-4260。doi:10.1091 / mbc.e07-04-0309。
36. Lin MT,等。Cyr61的表达升高通过整合素alpha2beta1增强胃癌细胞的腹膜扩散。J.Biol。化学 2007年;282:34594–34604。doi:10.1074 / jbc.M706600200。[ PubMed ] [ CrossRef ] [ Google Scholar ]
37.胡Q,郭C,李Y,Aronow BJ,张J。LMO7介导Rho-心肌素相关转录因子-血清反应因子途径的细胞特异性活化,并在乳腺癌细胞迁移中起重要作用。大声笑 细胞生物学。2011; 31:3223-3240。doi:10.1128 / MCB.01365-10。
38.施密德MC,等人。整联蛋白-alpha4beta1加细胞因子SDF-1alpha或IL-1beta的联合封锁有效地抑制了肿瘤的炎症和生长。癌症研究。2011; 71:6965-6975。doi:10.1158 / 0008-5472.CAN-11-0588。
39. Vlahakis NE等。整合素alpha9beta1直接与血管内皮生长因子(VEGF)-A结合,并有助于VEGF-A诱导的血管生成。J.Biol。化学 2007年;282:15187-15196。doi:10.1074 / jbc.M609323200。
40. Oneyama C,等人。MicroRNA介导的整合素连接激酶的上调促进Src诱导的肿瘤进展。癌基因。2012; 31:1623-1635。doi:10.1038 / onc.2011.367。
41. Danussi C等人。EMILIN1-alpha4 / alpha9整合素相互作用抑制皮肤成纤维细胞和角质形成细胞的增殖。J.细胞生物学。2011; 195:131–145。doi:10.1083 / jcb.201008013。
42.卓Y,查玛斯R,贝利斯SL。β1整合素的唾液酸化可阻止细胞粘附于galectin-3,并保护细胞免受galectin-3诱导的细胞凋亡。J.Biol。化学 2008年;283:22177-22185。doi:10.1074 / jbc.M800015200。
43. Gupta SK,Oommen S,Aubry MC,Williams BP,Vlahakis NE。整合素alpha9beta1通过增强上皮-间质转化促进恶性肿瘤的生长和转移。癌基因。2013; 32:141–150。doi:10.1038 / onc.2012.41。
44.郝建民,等。五基因签名可作为大肠癌转移和生存的潜在预测指标。J.Pathol。2010; 220:475-489。
45. Laplante M,萨巴蒂尼DM。mTOR信号一目了然。J.细胞科学。2009; 122:3589-3594。doi:10.1242 / jcs.051011。
46. Woodruff PG等。吸烟引起的独特的肺泡巨噬细胞活化状态。是。J.呼吸。暴击 护理医学。2005; 172:1383-1392。doi:10.1164 / rccm.200505-686OC。 [ PMC免费文章] [ PubMed ] [ CrossRef ] [ Google学术搜索]
点击查看:文章上部分总结内容
免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。
来源于:pubmed.nih
随机推荐
- 如何把图片中的英文翻译成中文?
- 什么是人工翻译?人工翻译怎么使用?
- 论文翻译怎么翻译精确?文档翻译超全教程
- 巴菲特致股东的封(2020)上
- 全文翻译:一键翻译整篇pdf文档
- ppt资料翻译用什么好?人工翻译怎么收费?
- 怎么在线图片翻译文字?翻译图片
- 郑州文档翻译平台哪个好?需要注意什么
- 武汉文档翻译平台哪个好?它有什么优势?
- 苏州免费在线翻译平台推荐以及有何优势
- 沈阳pdf翻译平台免费哪个好?哪个翻译平台靠谱?
- 在线翻译文档免费真的吗?在线翻译文档免费可靠吗?
- 三门峡文档翻译软件在翻译英文资料的时候要注意什么细节?医学翻译费用是多少钱?
- 太和文档英文翻译软件靠谱有哪个?怎么翻译文档?
- 永新word文档翻译如何操作?word文档翻译的简单方法是什么?