

 证件证明翻译
证件证明翻译 医疗病历翻译
医疗病历翻译 法律合同翻译
法律合同翻译 公司介绍翻译
公司介绍翻译 学术论文翻译
学术论文翻译 留学移民翻译
留学移民翻译 求职简历翻译
求职简历翻译 产品说明书翻译
产品说明书翻译
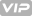




得益于分子胶,雄性果蝇的单个X染色体可以与雌性动物的两个X染色体一样活跃。
来源于:mpg
2020年11月18日
雄果蝇只有一个X染色体,而不像雌虫那样只有两个。因此,它必须具有两倍的活性才能达到相同的“基因剂量”。如果那不起作用,那么动物就会死亡。弗赖堡马克斯·普朗克免疫生物学和表观遗传学研究所的研究人员现在发现,负责所谓的剂量补偿的MSL复合物如何能够将X染色体与其他常染色体区分开。Asifa Akhtar的团队在小鼠中创建了微型蝇染色体,并鉴定了识别X染色体必不可少的分子成分。研究表明,利用X染色体合成的MSL2蛋白和roX RNA,两个组分必须一起形成一种凝胶。
MSL2和仅由雄性X染色体合成的roX2 RNA形成了一种凝胶状的胶粘剂(绿色凝胶),可将果蝇MSL复合体特异性结合到果蝇的X染色体上。
©Shutterstock:Anusorn Nakdee和Holiday.Photo.Top; 大会:弗莱堡免疫生物学和表观遗传学MPI
女性拥有X染色体的两个副本,具有1000多个基因。另一方面,男人只有一个拷贝,而有一个低基因的Y染色体。在动物界的许多物种中也发现了这种染色体失衡。但是,“平衡”这些差异至关重要。例如,雌性只关闭人类和小鼠的X染色体之一,果蝇则接管了果蝇。男性负责工作。一种称为MSL复合物的表观遗传因子与单个雄性X染色体结合,并利用其组蛋白乙酰化功能来过度激活X染色体。通过这种方式,它试图达到与雌性携带的两个X染色体相同水平的RNA产生。如果此过程失败,则雄蝇死亡。
“它使研究人员惊讶于MSL复合体如何知道八个飞行染色体中的哪个是X染色体,”弗莱堡马克斯·普朗克免疫生物学和表观遗传学研究所所长Asifa Akhtar解释说。这个问题促使她的团队设计出一种新颖而复杂的策略来研究MSL复合体如何识别X染色体。研究人员决定直接将MSL复合物移植到完全陌生的环境中:小鼠,而不是直接研究MSL复合物。
小鼠的微型X染色体染色体
Asifa Akhtar的团队能够通过在小鼠细胞中添加来自果蝇的MSL2和roX RNA来重建微型果蝇X染色体。使它们可见(右),并类似于在自然条件下进行剂量补偿时在雄蝇中也观察到的X染色体区域(由MSL复合物覆盖)。©免疫生物学和表观遗传学MPI / Keller-Valsecchi等。
不寻常的研究方法的一部分是回到基础上,并在小鼠中逐个分量地重建蝇染色体检测机制。该团队首先在小鼠中表达果蝇的MSL复合体MSL2中的一种蛋白质。起初他们看不到任何效果。但是基于对苍蝇的先前研究,他们怀疑可能需要MSL复合体的其他组件:非编码RNA分子roX1和roX2。现在,研究人员在为小鼠细胞动态配置了MSL2和roX2 RNA之后,现在清楚地观察到,在细胞核区域内,roX2以及涉及MSL2和roX2的点的基因活性都有所增加。
这些区域的特殊性质及其激活基因的潜力,非常让人联想到雄性蝇中由MSL复合体标记的X染色体上的区域。令人着迷的是,这些实验表明,在小鼠细胞中添加MSL2和roX2似乎足以在小鼠细胞中重建微型fly X染色体。这种创新方法强调了MSL复合体识别和激活果蝇X染色体第一步所需的最小分子成分。
鉴定X染色体的分子胶
现在,研究小组已经找到了形成微型fly X染色体的确切方法,他们将试管中的roX-RNA和MSL2两种成分混合在一起,以了解有关分子机制的更多信息。“当我们混合MSL2和roX RNA时,我们发现了一些有趣的东西。该研究的第一作者之一克劳迪娅•凯勒-瓦尔塞基(Claudia Keller-Valsecchi)说,这两种成分虽然都是孤立的液体,但它们开始形成球形颗粒,并变成了另一种看起来像凝胶的形状。有趣的是,RNA分子roX1和roX2均由X染色体上的基因编码,并且基因组中的该位置被用作MSL复合体组装的结合平台。
研究人员现在怀疑,从X染色体合成的roX RNA会“捕获”附近的MSL2分子,这是因为它们具有相互作用的潜力,并且倾向于变成凝胶状。“存在的roX RNA的量很好地指示了MSL复合物可以找到X染色体的程度。当我们在实验中改变roX RNA的数量时,从X染色体合成的roX越多,MSL复合物就能将X染色体与常染色体区分开来越好。合著者费利西亚·巴斯利卡塔(Felicia Basilicata)。
弗赖堡大学的科学家们凭借他们的研究结果,发现了一种新机制,该机制被雄蝇用来区分和标记单个X染色体,并且是基于roX-MSL2凝胶由两种成分组装而成的。无法将这种凝胶混合在一起的雄蝇会死亡。“也可以想到,凝胶状态可以帮助吸引和捕获其他重要成分以进行剂量补偿,例如转录机制,这对于增加RNA的产生是必需的。” Asifa Akhtar解释说,未来的研究问题仍然需要解决适用。
点击:查看医学类文章
其它分类文章
免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息,仅代表作者个人观点,与本网无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。
2020-11-23 18:21:01
科学家们确定了34,000岁的东亚早期混合欧亚血统尼安德特人在一项新的研究中,德国马克斯·普朗克进化人类学研究所和蒙古科学院的研究人员对迄今在蒙古发现的最古老的人类化石进行了基因组分析。他们表明,这名34,000岁的女性从西方欧亚人那里继承了大约25%的DNA,表明人们在欧亚大陆首次由当今人口的祖先定居后不久便迁移到整个欧亚大陆。这项研究还表明,这个人以及一个来自中国的40,000岁的人都携带了Denisovans的DNA,Denisovans是人类在人类到来之前就已居住在亚洲的一种灭绝的人类激素。在蒙古东部的索尔基特山谷发现的黄cap属于一个活在34,000年前的女性。分析表明:她从西方欧亚大陆继承了大约25%的DNA。 ©蒙古科学院考古研究所2006年,矿工在蒙古东部诺罗夫林县的索尔基特河谷中发现了具有特殊形态特征的人均黄skull。它最初被称为Mongolanthropus,被认为是穴居人,甚至是直立人。“ Salkhit”个体的遗体代表了该国唯一的更新世人化石化石。从无边便帽中提取的古代DNA显示,它属于一个生活在34,000年前的现代女性女性,与亚洲人比与欧洲人的关系更多。与迄今为止唯一的其他东亚早期个体进行基因研究的比较表明,这是一个来自中国北京郊外天元洞的40,000岁男性,这两个个体彼此相关。但是,它们之间的区别在于,萨尔基特人的祖先有四分之一来自西方的欧亚大陆,可能是通过与古代西伯利亚人混合而成的。迁移与互动研究人员Diyendo Massilani分析了Salkhit和Tianyuan个体的基因组,并发现了灭绝的人类激素的DNA痕迹。 ©MPI f。进化人类学马克思·普朗克进化人类学研究所的主要作者,研究者迪扬多·马西拉尼说:“这直接证明东亚的现代人类社区早在34,000年前就已经具有国际化性。” “这一罕见的标本表明,大约35,000年前,欧亚大陆各地人口之间的迁移和互动已经频繁发生”。研究人员使用了马克斯-普朗克进化人类学研究所开发的一种新方法,从萨尔基特和天元基因组中已灭绝的人类素中找到DNA片段。他们发现,这两个基因组不仅包含尼安德特人的DNA,而且还包含来自Denisovans的DNA,Denisovans是一个难以捉摸的亚洲尼安德特人的亲戚。“令人着迷的是,我们已经能够从中获得遗传数据的东亚最古老人类的祖先已经与Denisovans混合在一起,Denisovans是一种已灭绝的人类激素,已为当今亚洲和大洋洲的人口做出了贡献”,蒙古科学院考古研究所研究员Byambaa Gunchinsuren说。“这是Denisovans和现代人类在4万多年前相遇和融合的直接证据”。有趣的是,这些非常古老的东亚人中的Denisovan DNA片段与东亚当今人口基因组中的Denisovan DNA片段重叠,但在大洋洲中却与Denisovan DNA片段不重叠。这支持了Denisovans和现代人类之间多重独立混合事件的模型。” Massilani说。 点击:查看其他分类文章 查看天文学文章 使用文档翻译功能免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息,仅代表作者个人观点,与本网无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源:MPG
2020-11-18 19:31:50
具有脂质特异性寡克隆IgM谱带特征的患者外周血单个核细胞的全转录组分析揭示了两个环状RNA和两个线性RNA作为高活性疾病的生物标志物患者核细胞揭示环状线性RNA高活性疾病生物标志物(上)3.3. 线性成绩单表达谱在本研究中,我们还通过RNA序列分析了线性转录组,鉴定出115,869个转录本,总读取次数≥10。我们使用检测标准应用了一个额外的过滤器,该过滤器可以识别出具有一致表达模式的84,863个线性RNA,从两组中均检测到92.8%。其中,有2441个转录物被差异表达(p<0.05和FC> | 2 |),两组均检测到1421个转录物(图3B)。其余的转录本是特定于一组的,仅在LS-OCMBs阴性患者的PBMC中表达382份,而在LS-OCMBs阳性的患者中表达638份。我们选择了十个候选线性RNA来通过RT-qPCR确认它们的差异表达。更大的队列。如图3D所示,在LS-OCMB阳性患者的PBMC中证实了IRF5和MTRNR2L8的较低表达(两个转录本中FC = -1.33; p <0.05)。差异表达的线性RNA主要在免疫系统的生物过程中富集。图4显示了最有意义和最丰富的术语(FDR<0.01和倍富集> 2)。补体激活,体液免疫应答和I型干扰素信号传导路径出现在最丰富的术语中。 图4. 2441 DEmRNA的基因超表达测试结果。显示了最显着的(倍富集> 2)和富集的(FDR <0.01)GO生物过程。条形根据其FDR值进行着色。 DE-差异表达。 FDR-错误发现率。3.4. circRNA和线性RNA作为高度活跃疾病的生物标志物的评估为了评估四个经过验证的RNA作为LS-OCMB状态血液生物标志物的潜力,我们进行了ROC曲线分析。如表2所示,测试了不同的RNA组合以找到区分两组的最佳性能。我们发现,circRNA和线性RNA的不同组合可改善性能,达到约70%的AUC值。表2.四个候选成绩单和不同组合的ROC分析结果。ROC-接收机的工作特性。4. 讨论区在这项工作中,我们表征了具有明显LS-OCMB状态的MS患者的PBMC的整个转录组。该分析和验证实验表明,LS-OCMB阳性患者的PBMC的整体转录组与LS-OCMB阴性患者的PBMC的整体转录组不同。RNA-seq结果显示,两组之间有124个circRNA和2441个线性RNA差异表达。有趣的是,仅在一组中检测到58%的线性RNA,突显了来自具有不同LS-OCMB状态的患者的PBMC中存在特定的表达模式。是真的;但是,我们的RNA-seq样本数量有限,应谨慎对待这些观察结果。据我们所知,以前没有circRNA与MS相关。 Circ_0000478位于13号染色体,其宿主基因为von Willebrand因子A结构域,包含8个(VWA8)。有趣的是,该基因的线性转录本也是我们数据集中差异表达的线性RNA之一,其折叠变化为-3.23(p = 0.047)。最近已经描述了该转录本,并编码功能仍不确定的线粒体蛋白[32]。关于circ_0116639,它位于22号染色体上,与EP300基因重叠。 EP300是一种组蛋白乙酰转移酶,在我们的研究中被检测到但没有差异表达。 circRNA及其宿主基因表达模式的这些差异揭示了circRNA的生物发生与其宿主基因的生物发生是独立调控的,如先前所述[33]。值得注意的是,干扰素调节因子5(IRF5)是已确认的下调转录物之一,该基因的多态性与国际多发性硬化症遗传学联盟进行的最后一次全基因组关联分析中存在发展MS的风险有关。以及西班牙人群的复制研究[34,35]。此外,该基因中的SNP与CSF中CXCL13的水平增加有关,CSF是与高度活跃的疾病相关的趋化因子[36]。最重要的是,动物模型显示IRF5与小胶质细胞极化有关对中风和阿尔茨海默氏病有炎性反应(M1)[37,38]。正如其他作者在外显子组测序项目中所暗示的那样,这可能暗示了小胶质细胞在更积极的疾病过程中的意义[39]。尽管如此,我们的发现是在外周PBMC中进行的,因此IRF5来源可能是外周单核细胞或巨噬细胞。无论如何,需要进一步的研究来揭示IRF5在高度活跃的MS疾病病程中的意义。另一方面,MT-RNR2像8转录本(MTRNR2L8)是另一种经过验证的线性转录本,在LS-OCMB+组中表达较低,编码在11号染色体上,有趣的是,它跨越了miR-4485茎的位置环定位,是本研究中选择用于验证的四个miRNA之一。根据微阵列数据,miR-4485在MTRNR2L8中被下调在LS-OCMB +组中。 MTRNR2L8是mir-4485的宿主基因这一事实可能解释了,即使无法验证miRNA,这组患者的两个转录本也都被下调了。这些结果表明这两个RNA之间可能存在相互作用,可能与更活跃的疾病有关,但是需要进一步的研究来证实这一假设。根据UniProt的研究,MTRNR2L8具有神经保护和抗凋亡因子的作用,并且发现重度抑郁症患者在特定区域的大脑中MTRNR2L8的含量增加[40]。尽管该基因的功能尚不清楚,但对于MS而言,作为神经保护因子的可能作用非常有趣,因为我们发现该基因在PBMC中被下调。但是同样,应该进行进一步的研究以揭示外周血中MTRNR2L8表达与疾病活动以及这种关系的潜在机制之间的可能联系。基因过度表达测试表明,与补体激活,体液免疫反应和I型干扰素反应有关的生物学过程是最丰富的术语。值得注意的是,IgM抗体的鞘内合成以前与鞘内补体激活相关[41],并且已经清楚地证明了其在脱髓鞘和轴突损伤中的作用[42]。有趣的是,补体系统的不同组成部分已与MS相关,某些组成部分的血浆水平已被提议作为MS疾病状态生物标志物[43]。值得注意的是,这些过程在具有高活性疾病标志物的患者的PBMC中的改变基因中富集。该分析可能表明这些患者的免疫反应可能改变,这可能解释了其较高的疾病活动性,但仍需要进一步和其他类型的研究来探索这一假设。这项研究的主要目的是鉴定血液中的一种或多种可以区分阳性和阴性LS-OCMB患者的生物标志物,从而可以用作高度活跃疾病的更容易获得的标志物。 ROC曲线分析显示,这些标记组合在一起具有大约70%的准确度,通常被认为是可以接受的性能[44]。考虑到这些都是在血液中测量的,它们可能有助于诊所进行重复测量,而连续采集CSF样品则存在严重缺陷。鉴于他们的有效治疗范围可能比较良性或较不活跃的疾病形式要窄,因此正在努力确定患有高度活跃疾病的患者[45]。与此相一致,已经提出了几种生物标记物来鉴定具有侵略性疾病进程的个体,例如神经丝轻链的CSF水平,CXCL13或CHI3L1 [46-48]。其他作者还报告了具有不同疾病活动的患者之间非编码转录组的差异。 Quintana及其同事研究了一组具有LS-OCMB特征的MS患者和其他神经系统疾病(OND)患者的miRNA在CSF中的表达[49]。他们发现OND组和LS-OCMB +组之间miR-30a-5p,miR-150,miR-645和miR-191的表达存在差异,但他们发现LS-OCB阳性和阴性患者之间没有任何差异OCMB。由于他们的研究与本研究之间的分析平台不同,我们无法检查来自Quintana及其同事的数据中候选miRNA的表达。另一项研究报道,miR-24-3p的血清水平与疾病进展指数相关,通过计算EDSS值除以疾病持续时间得出,但未测量LS-OCMB的状态[50]。最近,长非编码RNA在血液中的表达也被描述为显示根据年龄相关的MS严重程度评分,区分高活跃MS患者与其他病程较轻的患者的能力[51]。所有这些研究都突出了能够识别高活动性疾病患者的生物标志物的需求,以及科学界朝着这个方向所做的努力。此外,鉴于每项研究使用不同的参数来对各组患者进行测量和分类,因此对于什么是侵略性疾病过程的定义达成共识也很重要,这使得它们之间的比较确实具有挑战性[52 ,53]。该组患者的早期识别可以受益于不同的治疗建议,并且;因此,有可能改善疾病的长期结果。我们的miRNA分析研究发现了两组之间差异表达的miRNA,尽管尚未通过RT-qPCR实验验证。在这方面,其他技术(例如液滴数字PCR(ddPCR))可能有助于减少变异性并发现组之间的细微差异。尽管据报道在基因表达研究中使用ddPCR并不会增加灵敏度,并且只要起始物质的量足够多且反应中没有污染物,其性能是相似的[54,55]。尽管如此,在未来的验证项目中可以考虑使用该技术,该项目旨在明确丢弃或将那些miRNA用作生物标志物,以用于更具侵略性的MS疗程。5. 结论总之,我们已经鉴定出在LS-OCMB阳性患者中被下调的两个circRNA和两个线性RNA。我们的发现对于多发性硬化症的生物标志物领域非常重要,因为它们可以有助于识别高度活跃的疾病患者。此外,与其他生物标志物相比,一个重要的优势是可以在血液中检测到它们,从而可以进行连续检测来监测其水平,而腰椎穿刺不建议这样做。为了确定其效用,还需要在更大的人群中进行进一步的研究,但是我们认为,它们可以作为确定哪些患者应进行腰穿以确认其LS-OCMB状态的首选筛查工具。 补充材料:补充材料可以在http://www.mdpi.com/2227-9059/8/12/540/s1上找到。表S1:样品和实验的完整列表,其中包括每个样品。表S2:RT-qPCR实验中使用的测定和引物。图S1:PCR产物的琼脂糖凝胶电泳图像和Sanger测序结果显示circRNA的反向剪接。作者贡献:概念化,D.O。 (DavidOtaegui)和M.M.-C .;方法论(DavidOtaegui),M.M.-C.,L.I.,TB.D.O。(Danel Olaverri)和L.M.V .;验证和L.S .;形式分析,L.I.,D.O.(DanelOlaverri),TB,M.M.-C.,M.E。和L.C.-F .;调查,L.I.,M.M.-C.,M.E.,L.C.-F .;资源,A.P.,T.C.-T.,M.E.,L.C.F.,L.M.V。和D.O. (David Otaegui);写作-原始草稿,L.I.,D.O。 (David Otaegui)和M.M.-C .;写作-审查和编辑(Danel Olaverri),TB。,Á.P.,TC.C.T.,L.M.V .;可视化,L.I.,D.O. (Danel Olaverri),M.M.-C .;监督,D.O.(DavidOtaegui),美国密西西比州立大学和L.M.V .;项目管理,做。 (David Otaegui)和M.M.-C .;资金收购,L.M.V.,D.O. (David Otaegui)和M.M.C.所有作者均已阅读并同意该手稿的发行版本。资金:这项研究由萨洛德·卡洛斯三世研究所(PI17 / 00189和PI15 /00513),西班牙红色硬化菌(REEM)(RD16/0015/0007和RD16/ 0015/0001),吉普斯夸省理事会(109)资助/ 18)和巴斯克政府(PRE_2019_2_0206)和ELKARTEK计划。致谢:作者要感谢Biodonostia Health Research Institute基因组平台的工作人员。利益冲突:作者声明没有利益冲突。资助者在研究的设计中没有作用。在数据的收集,分析或解释中;在手稿的撰写中,或在决定发表结果时。 参考文献(展示部分文献内容,查看全部可在原网站)1. 菲利皮,M .;Bar-Or,A。 Piehl,F .; Preziosa,P .; Solari,A .; Vukusic,S .;马萨诸塞州罗卡多发性硬化症。纳特版本号Dis。总理。 2018,4,1–27。[CrossRef] [PubMed]2. M.Comabella; X. Montalban。多发性硬化症中的体液生物标志物。柳叶刀神经。 2014,13,113–126。 [CrossRef]3. 保罗M.Comabella; Gandhi,R.多发性硬化症中的生物标志物。冷泉湾。透视。中2019年9. [CrossRef] [PubMed]4. 维拉尔(L.M.);马里兰州萨达巴; E. Roldan; Masjuan,J .;冈萨雷斯-波克(P.); N.比利亚鲁比亚;马萨诸塞州埃斯皮诺;加西亚·特鲁希略(J.A.); Bootello,A .; Alvarez-Cermeno,J.C.鞘内合成针对髓磷脂的寡克隆IgM预测了MS的侵袭性病程。 J.临床。调查。 2005,115,187–194。 [CrossRef]5. 小波利夫卡(J.J.P.);克拉科罗娃;彼得卡,M。 O.Topolcan。神经病学中生物标志物研究的现状。EPMA J.,2016年,第1-13页。[CrossRef]6. Kemppinen,A.K .;卡普里奥(Kaprio)帕洛蒂(Palotie) Saarela,J.多发性硬化症中全基因组表达研究的系统评价。 BMJ Open 2011,1,e000053。 [CrossRef]7. 尼克斯;陈慧平;李明Khankhanian,P .; Madireddy,L .; Caillier,S.J .; A. Santaniello;克里(Cree)佩莱蒂埃(D.) Hauser,S.L .;等。大量多发性硬化症患者和健康对照人群的血液RNA分析。哼。大声笑基因2013,22,4194-4205。 [CrossRef] [PubMed]8. Irizar,H .; Muñoz-Culla,M .; Sáenz-Cuesta,M。 Osorio-Querejeta,I .;塞普尔韦达(L.卡斯蒂略-特里维尼奥(T.普拉达(A.) Lopez de Munain,A.;J.Olascoaga;Otaegui,D.通过差异性ncRNA-mRNA网络分析将ncRNA鉴定为多发性硬化症的潜在治疗靶标。 BMC Genom。 2015,16,250. [CrossRef] [PubMed]9. S.Dolati;马洛菲Z.Babaloo;阿格巴蒂-马累基罗珊加湖M.Ahmadi;里赫特加河;Yousefi,M.与多发性硬化症发病机制有关的miRNA网络失调。生物医学。药剂师。2018,104,280–290。 [CrossRef]10. 杜C.刘超康建赵庚;是的,Z。黄胜李正吴中;裴G;杜C.等。 MicroRNA miR-326调节TH -17分化,并与多发性硬化症的发病机制有关。纳特免疫2009,1259,1252-1259。[CrossRef] [PubMed]11. Muñoz-Culla,M .; Irizar,H .; Sáenz-Cuesta,M。卡斯蒂略-特里维尼奥(T. Osorio-Querejeta,I .;塞普尔韦达(L. LópezDeMunain,A .;J.Olascoaga; Otaegui,D.在多发性硬化症复发和缓解中发现的SncRNA(microRNA&snoRNA)相反表达模式是性别依赖性的。科学Rep。2016,6. [CrossRef]12. DeFelice,B.;蒙多拉A.萨索;Orefice,G.;布雷西亚莫拉瓦卡,G。 E. Biffali;M .; Pannone,R.用干扰素-β治疗后多发性硬化症患者的小非编码RNA信号。 BMC Med。基因组2014,7,1–9。[CrossRef][PubMed]13. 乔纳斯(Jonas) Ezaurralde,E.对microRNA介导的基因沉默的分子理解。纳特吉内特牧师2015,16,421–433。 [CrossRef]14. 弗洛里斯,G。张丽;Follesa,P .; Sun,T.环状RNA和神经系统疾病的调节作用。大声笑神经生物学。 2017,54,5156-5165。 [CrossRef]15. 夏X.唐X. Wang,S.CircRNA在自身免疫性疾病中的作用。面前。免疫2019,10,1–8。 [CrossRef] [PubMed]16. E.M. Paraboschi;Cardamone,G.;索尔达,G .; Duga,S.; Asselta,R.解释多发性硬化症基因组范围相关区域的非编码遗传变异。面前。基因2018,9,1-10。 [CrossRef]17. Cardamone,G .; E.M. Paraboschi; Rimoldi,V .;Duga,S.;索尔达,G .; Asselta,R. GSDMB剪接和反向剪接图谱的表征鉴定了在多发性硬化症中失调的新型同工型和环状RNA。诠释J.摩尔科学2017,18,576。[CrossRef]18. 伊帕拉吉雷(L. Muñoz-Culla,M .;普拉达-伦戈,I .;卡斯蒂略-特里维尼奥(T. J.Olascoaga; Otaegui,D.环状RNA分析显示ANXA2的环状RNA可用作多发性硬化症的新生物标记。哼。大声笑基因2017,26. [CrossRef]19. M.J. Kacperska;Walenczak,J。Tomasik,B.血浆microRNA作为多发性硬化症的潜在生物标志物:文献综述。进阶临床经验中2016,25,775–779。 [CrossRef]20. Stoicea,N。杜阿哥伦比亚特区拉基斯;蒂普顿角;阿里亚斯-莫拉雷斯(C.E.); Bergese,S.D.从理论到实践作为CNS生物标记物的MiRNA之旅。面前。基因2016,7,1–8。 [CrossRef]21. 张正。杨涛; Xiao,J.环状RNA:人类疾病的有前途的生物标志物。 EBioMedicine 2018,34,267–274。 [CrossRef][PubMed]22. 维拉尔(L.M.); González-Porqué,体育;Masjuán,J.; Álvarez-Cermeño,J. Bootello,A .; Keir,G.一种检测寡克隆IgM条带的灵敏且可重复的方法。 J.免疫。方法2001,258,151–155。 [CrossRef]23. 肯特(W.J.); C.W. Sugnet; T.S. Furey;罗斯金(K.M.);普林格尔(T.H.); Zahler,A.M.; Haussler,A.D. UCSC的人类基因组浏览器。基因组研究。2002,12,996–1006。 [CrossRef] [PubMed]24. A.Dobin;戴维斯Schlesinger,F。 J.Drenkow; Zaleski,C。 Jha,S.;巴图,P.;Chaisson,M.; Gingeras,T.R. STAR:超快速通用RNA序列比对仪。生物信息学2013,29,15–21。 [CrossRef]25. 李华;Durbin,R.与Burrows-Wheeler变换快速准确地进行短读比对。生物信息学2009,25,1754–1760。 [CrossRef]26. 张旭。董河;张Y张建。罗Z.张建。陈力; Yang,L.环状RNA的多样交替反向剪接和交替剪接。基因组研究。 2016,26,1277–1287。[CrossRef]27. 高Y张建。Zhao,F.基于多重种子匹配的环形RNA鉴定。简要。生物信息。2017,1–8。 [CrossRef]28. 汉森(T.B.通过结合预测算法改进的circRNA鉴定。面前。单元开发。生物学2018,6,1–9。 [CrossRef]29. 爱,密歇根州;休伯,W。Anders,S.使用DESeq2对RNA-seq数据的倍数变化和分散度进行适度估计。基因组生物学。 2014年15月1日至21日。 [CrossRef]30. N.L.布雷; Pimentel,H .; P. Pachter,L.接近最佳概率的RNA序列定量。纳特生物技术。 2016,34,525–528。[CrossRef]31. Mi,H .; Thomas,P。PANTHER Pathway:基于本体的路径数据库,结合了数据分析工具。纳特协议。 2019,14,703–721。 [CrossRef] [PubMed]32. 罗,M。马威;沙,Z。 J.Finlayson。王婷; Diaz,R.;威利斯(W.T.);Mandarino,L.J.Von Willebrand因子A结构域含蛋白8(VWA8)定位于线粒体内膜的基质侧。生化。生物物理学。 Res。社区2020,521,158–163。[CrossRef]33. 摩尔多瓦,L.I .;汉森,TB。威尼托; Okholm,T.L.H .; T.L.安德森; Hager,H .;艾弗森,L。 Kjems,J .; Johansen,C .;克里斯滕森(L.S.)成对的病变和非病变皮肤的高通量RNA测序揭示了牛皮癣circRNAome中的主要变化。 BMC Med。基因组2019,12,1-17。 [CrossRef] [PubMed]34. 国际多发性硬化症遗传学联盟。多发性硬化症基因组图谱暗示易感性周围免疫细胞和小胶质细胞。 Science 2019,365,1-10。 [CrossRef]点击:查看更多生物学文章 使用文档翻译功能免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:mdpi
2020-11-18 19:00:00
每年,世界各地的人们都遭受约1.8-270万蛇毒咬伤。由于无法获得可用的治疗方法或健康保险,许多此类伤害得不到治疗,导致10万多人死亡,约是永久性残疾的三倍。需要新的疗法来治疗蛇咬伤,这是被忽视的公共卫生问题。一种新的治疗蛇毒咬伤的治疗工具可能来自不太可能的来源-干细胞研究。 在非洲,亚洲和拉丁美洲,约有250种蛇毒被蛇毒咬伤的患病率最高。蛇毒由多种天然毒素,酶,蛋白质和肽组成,它们共同起作用,迅速引起组织损伤或死亡。确切的毒液成分极其多样且取决于物种,因此对每种蛇都具有高度特异性。蛇咬伤可致死或导致终身慢性健康问题,例如神经毒性和神经相关性麻痹,肾脏过度出血后不可逆的肾脏损害或肢体截肢。当前大多数的治疗旨在中和毒液,例如抗蛇毒子。通常,抗蛇毒素是通过人工挤蛇来收获蛇毒的方法(参见图1),并将小剂量的毒液注入供体动物(如马)中以引发免疫反应和产生抗体。然后将这些抗蛇毒抗体注射到蛇咬受害者中,在其中它们选择性地识别并结合蛇蛇毒的结构成分,从而中和毒素。 以这种方式制作抗蛇毒液非常耗时,难以扩大规模,并且必须针对每种蛇毒蛇单独进行。根据世界卫生组织的资料,在过去的20年中,许多抗蛇毒毒素生产商已经停止生产,从而导致供应减少和价格急剧上涨。不幸的是,缺乏有效的替代治疗方法。使用干细胞对抗蛇咬伤 最近,科学家转向干细胞来解决这个问题。研究人员可以使用干细胞来生长称为器官的三维微型器官,该器官可以维持特定器官或组织的基本特征。科学家们第一次尝试从蛇干细胞中提取类器官,以观察它们是否可以在实验室中生长出产生毒液的微型器官。研究人员解剖了9种蛇的毒腺,并成功地从蛇唾液腺中发现的干细胞中生长出类器官(见右图2)。图2:来自毒蜘蛛的蛇毒腺类器官。图片来源:拉维安·范因文(Ravian van Ineveld),荷兰马克西玛公主小儿肿瘤中心。令人惊讶的是,这些类器官包含的细胞可以分泌功能活跃的毒液,这种毒液会对实验室的神经元和肌肉细胞产生特定种类的作用。此外,类器官来源的毒液的成分与直接从蛇中提取的毒液相似。现在,科学家们希望为所有与医学有关的有毒蛇建立一个蛇体类生物体的生物库或储存库,以创造无限量的毒液。自发表最初的研究论文以来,Jens Puschhof说,该组织已经将类器官库扩大到包括更多种类的毒蛇。他补充说,使用类器官的毒液代替挤奶蛇可以帮助生产更安全,更可靠的产品,因为个别蛇的毒液成分存在差异。下一步包括在实验室而不是在马匹中使用毒类类器官来纯化有毒成分并制造抗体,以更有效地制造解毒剂并增强全球通行能力。 蛇毒能治疗肿瘤吗? 蛇毒类器官生物库可能会为其他疾病提供治疗。美国食品药品监督管理局(FDA)先前已批准从蛇毒中提取药物,用于治疗急性脑梗塞,急性冠状动脉综合征和预防手术中的出血。令人惊讶的是,早期研究表明,蛇毒中的肽可能具有抗肿瘤作用。蛇毒称为整合素,如Contortrostatin的部件从蛇铜头蝮contortrix,能抑制在人转移性皮肤癌的实验室模型的血管形成和癌细胞的粘附性。其他研究表明,在毒液中发现的有效肽可能会干扰血管的形成,从而减少乳腺癌小鼠模型中营养物质向肿瘤的传递。毒液生物库的可用性将使科学家能够进一步研究蛇毒成分,以查看是否存在生物医学创新的机会。咬每五分钟有50人被蛇咬伤,在这场默默的医疗危机中,有一个人会死。新颖的干细胞研究已使科学家能够在培养皿中成功生长出产生微毒的类器官,可将其用于开发可及且负担得起的抗蛇毒血清疗法。开发蛇毒腺类有机体的这项科学创新有可能使全世界的人们免于致命的蛇咬和其他疾病。______________________________________________________点击:查看更多生物学文章 试用免费文档翻译 免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源:isscr
2020-11-17 19:11:29
来源:PHYS 罗格斯大学 Todd Bates着DNA双螺旋的三种构象:A(左),B(中)和左手Z(右)。图片提供:David S. Goodsell和RCSB PDB 罗格斯大学的科学家们认为,达尔文的进化理论应该扩展到包括考虑DNA稳定性的“能量代码”(所谓的“分子达尔文主义”),以进一步说明物种特征在地球上的长期生存。根据罗格斯(Rutgers)合着的研究,标志性遗传密码可被视为遵循热力学定律(能量流)演化的“能量密码”,导致其进化最终以几乎所有生物物种的奇异密码产生。发表在“生物物理学季刊”上。高级作者肯尼思·J·布雷斯劳尔,莱纳斯·鲍林杰出大学化学系教授和高级研究员肯尼斯·布雷斯劳尔说:“这些启示之所以重要,是因为它们提供了分析人类基因组和任何活着物种的基因组的全新方法,是生命的蓝图。”罗格斯大学-新不伦瑞克艺术与科学学院的化学生物学。他还隶属于新泽西州罗格斯癌症研究所。“ DNA遗传密码的进化起源和所有生物物种的进化都嵌入了其分子DNA蓝图的不同能量谱中。在热力学定律的影响下,这种能量密码从天文数字中演化而来。替代的可能性,转化为所有生物物种几乎相同的代码。”科学家们研究了这种所谓的“普遍之谜”,探究了惊人发现的起源,即遗传密码演变成几乎统一的蓝图,该蓝图源于数万亿种可能性。科学家将具有里程碑意义的“适者生存”达尔文进化论的基础扩展到了“分子达尔文主义”。达尔文的革命理论基于物种物理特征的世代持久性,这些特征使物种能够通过“自然选择”在给定的环境中生存。分子达尔文主义指代持续存在的物理特征,因为编码这些特征的分子DNA区域异常稳定。Breslauer说,不同的DNA区域可以表现出不同的能量特征,这些特征可能有利于能够实现特定生物学功能的生物体的物理结构。下一步包括将人类基因组化学序列重铸并绘制到“能量基因组”中,从而使具有不同能量稳定性的DNA区域与物理结构和生物学功能相关联。这将能够为基于分子的治疗方法更好地选择DNA靶标。 点击:查看更多生物学文章 查看更多医学文章 使用双语译文翻译免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。
2020-11-17 18:31:24
细菌可以通过复制另一个基因来补偿tRNA基因的丢失来源:MPG 2020年11月3日位于布隆(Plön)的马克斯·普朗克(Max Planck)进化生物学研究所的科学家利用实验室中细菌种群的实验性进化,直接观察到了tRNA基因的诞生。tRNA分子示意图。反密码子将密码子(在遗传信息中)与氨基酸(构成蛋白质)匹配。©MPI f。进化生物学 翻译是将遗传信息转化为蛋白质(细胞的主要力量)的过程。称为转移RNA(“ tRNA”)的小分子在翻译中起着至关重要的作用。它们是使密码子与氨基酸匹配的衔接子分子。生物携带多种类型的tRNA,每种均由一个或多个基因(“ tRNA基因集”)编码。广义上讲,tRNA基因集的功能(将61种密码子翻译成20种不同的氨基酸)在整个生物体中都是保守的。但是,tRNA基因集的组成在不同生物之间可能有很大差异。这些差异如何产生以及为什么产生,一直是科学家们长期关注的问题。 实验室中细菌tRNA集的进化 一个tRNA基因(红色X)的缺失可通过另一个tRNA基因(绿色)的复制来补偿。 马克斯·普朗克进化生物学研究所研究组负责人詹娜·加里(Jenna Gallie)和她的研究小组结合了数学建模和实验室实验,研究了荧光假单胞菌细菌的tRNA基因组如何进化。”我们首先从细菌基因组中去除一种类型的tRNA,从而导致细菌菌株缓慢生长。在实时进化实验中,我们为这种生长缓慢的菌株提供了改善其生长的机会。我们看到压力反复不断地改善。这种改善是由于大量细菌遗传信息的重复,每次重复都包含一个补偿性tRNA基因。最终,一种tRNA类型的消除通过增加另一种不同的tRNA类型的数量来补偿。” 珍娜·加莉(Jenna Gallie)说。复制的tRNA类型可以补偿,因为它能够以较低的速率执行消除的tRNA类型的密码子氨基酸匹配功能。相关基因组中tRNA基因的比较以前为整个进化史中某些tRNA基因的重复提供了证据。这里描述的实验提供了直接的经验证据,证明tRNA基因组可以通过复制事件进化。点击:查看更多生物学文章 查看更多医学文章 试用双语译翻译功能 免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息,仅代表作者个人观点,与本网无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。
2020-11-13 20:11:19
全球档案馆收集了三十年来的运动数据,记录了北极动物行为的变化2020年11月5日冬季更暖,春季更早,冰层不断缩小以及人类发展加快,北极正在经历着巨大的变化,正在影响着本土动物。现在,来自世界各地的研究人员已经建立了一个档案库,用于记录北极和亚北极地区动物的活动数据,该档案库位于德国拉多夫采尔的马克斯·普朗克动物行为研究所的Movebank平台上。借助北极动物运动档案馆,科学家们可以分享他们的知识,并协作提出有关动物如何应对不断变化的北极的问题。档案馆的三项最新研究揭示了该地区金鹰,熊,北美驯鹿,驼鹿和狼的行为的大规模模式,并说明了如何利用档案馆来识别更大的生态系统变化。 在一年中,龋齿会长距离迁移。版权©iStock / Geoffrey Reynaud长期以来,研究人员一直在观察北极动物的活动和行为。但是,很难发现和访问这些数据。为了解决这个问题,由Radolfzell的马克斯·普朗克动物行为研究所的数据策展人Sarah Davidson和俄亥俄州立大学的教授Gil Bohrer领导的国际团队已经建立了NASA资助的全球动物研究数据档案库在北极和亚北极地区迁移。北极动物运动档案馆的目的是与科学家建立联系并促进他们的合作。这特别重要,因为北极地区遍布世界各地。来自17个国家/地区的100所大学,政府机构和保护组织的研究人员参与了存档,该存档存放在由马克斯·普朗克研究所所长Martin Wikelski开发的研究平台Movebank上。Wikelski说:“我们的目标是利用档案馆建立跨越机构和政治边界的全球社区。” 该档案馆目前包含200多个项目,从1991年至今,已有8,000多种海洋和陆生动物的运动数据。金鹰的迁移方式 冬季过后,金鹰返回北极的繁殖地。©Adobe Stock / Tomas Hulik研究人员最近发表了一系列发现,利用档案库将数据和专业知识整合在一起。通过比较1993年至2017年100余只金鹰的运动,研究人员发现,春季迁徙到北方的未成熟鸟类在较温和的冬季之后更早到达。然而,成年鸡的到来时间一直保持恒定,无论其繁殖场的条件如何,都会影响到筑巢和雏鸡的生存。“黑岩森林”科学家斯科特·拉波特(Scott LaPoint)表示:“我们的方法揭示了评估跨越几代人和数十年气候模式的数据的重要性,如果忽略这些数据,它们会极大地影响我们的结果并进而影响管理策略。”对2000年至2017年间900多只北美驯鹿的第二项研究发现,更多的北方牧群在春季早些时候分娩,而更多南方种群的产犊日期并未显示出相同的变化。“在如此偏远和严酷的环境中,对于种群,亚种以及超过数百万平方公里的生物过程,如出生,进行如此大规模的观察,是前所未有的。这些结果揭示了我们不会怀疑的模式,并指出了从驯鹿进化到适应环境变化的能力等各方面的进一步研究。”马里兰大学的Elie Gurarie解释说。对气候变化的不同反应第三项分析研究了1998年至2019年熊,北美驯鹿,驼鹿和狼的运动速度,结果表明该物种对季节性温度和冬季降雪条件的反应不同。在华盛顿大学进行这项研究的彼得·马奥尼(Peter Mahoney)说:“动物如何通过运动应对变化的天气条件,将对物种竞争和捕食者-猎物的动态产生有趣的影响。”除了已经包含在档案中的数百项研究之外,随着数据从田间动物中传输以及越来越多的研究人员加入,资源也在不断增长。这应该有助于发现动物行为的变化,并最终发现整个北极生态系统的变化。戴维森说:“我们还为过去的行为和动作提供了急需的基线。” “这可用于改善野生动植物管理,解决关键的研究问题,并为子孙后代记录北极地区的变化”。点击:查看更多生物学文章 查看更多关于动物的文章 查看双语译文文章免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源:MPG
2020-11-13 19:26:31
随机推荐
- 如何把图片中的英文翻译成中文?
- 什么是人工翻译?人工翻译怎么使用?
- 论文翻译怎么翻译精确?文档翻译超全教程
- 巴菲特致股东的封(2020)上
- 全文翻译:一键翻译整篇pdf文档
- 洪泽pdf文档翻译哪个好?如何选择正规的翻译软件?
- 福州免费在线翻译哪个好?怎么操作?有什么优点?
- 绍兴pdf在线翻译用什么平台好?什么翻译平台靠谱?
- 免费的文档翻译软件真的存在吗?免费的文档翻译软件好用吗?
- 海口哪个中英翻译平台好用?翻译PDF文件该怎么操作?
- word文档翻译平台费用与使用流程
- 无需手动转换,有没有能翻译PDF的软件?
- 周宁在线翻译文档推荐哪个?在线翻译文档费用是多少?
- 伊春word翻译软件哪个好用?文献翻译的技巧有哪些?
- 昌平区文档翻译哪家好?翻译平台是怎么翻译文档的?