



 证件证明翻译
证件证明翻译
 医疗病历翻译
医疗病历翻译
 法律合同翻译
法律合同翻译
 公司介绍翻译
公司介绍翻译
 学术论文翻译
学术论文翻译
 留学移民翻译
留学移民翻译
 求职简历翻译
求职简历翻译
 产品说明书翻译
产品说明书翻译















 剩余额度:
剩余额度:
 个人中心
个人中心
 我的翻译
我的翻译
 优惠券
优惠券
 修改密码
修改密码
 退出登录
退出登录

SARS-CoV-2的颗粒(绿色;人工着色)感染宿主细胞。图片来源:AMI图片/科学图片库 3月10日-令人担忧的冠状病毒变种为更好的疫苗带来希望 被快速传播的冠状病毒变异感染的人会产生免疫反应,可以抵御多种SARS-CoV-2病毒株。 科学家于2020年末在南非首次发现了SARS-CoV-2变种,称为B.1.351。此后,他们将其与再感染相关联,并发现提示,几种疫苗对它的有效性不及对早些时候流行的SARS-CoV-2变种的影响。大流行。 南非约翰内斯堡国家传染病研究所的Penny Moore及其同事评估了89名感染B.1.351并入院的人的抗体反应(T. Moyo-Gwete等人,bioRxiv的预印本)https://doi.org/fzq5 ; 2021)。研究小组发现,这些参与者的抗体水平与早期菌株感染者的抗体水平相似。 然后,研究小组对感染了B.1.351的人的抗体进行了抗化处理,使其针对某种形式的HIV病毒进行了修饰,以使用冠状病毒刺突蛋白感染细胞。这些抗体能够灭活结合了B.1.351中发现的刺突蛋白形式的病毒,早期菌株以及巴西鉴定出的新兴变异体P.1。。 这组作者说,结果表明,基于B.1.351基因序列的疫苗可以保护人们免受多种冠状病毒的侵害。调查结果尚未经过同行评审。 实验表明,他的免疫应答称为T细胞(人为着色)可以抵御新的SARS-CoV-2变体。图片来源:Steve Gschmeissner /科学图片库 3月5日-T细胞可能会从猖cor的冠状病毒变异中拯救 实验室研究表明,新兴的冠状病毒变种似乎并未躲避重要的免疫系统角色,即T细胞。 最近发现的一些SARS-CoV-2变体可以部分规避因接种疫苗和先前感染而产生的抗体,这引发了人们的担忧,即针对变体的疫苗比针对原始病毒株的疫苗更无效。加利福尼亚拉霍亚免疫学研究所的Alessandro Sette和Alba Grifoni及其同事研究了这些变体的突变是否也可能帮助他们逃避了T细胞-免疫系统的组成部分,对于降低传染病的严重性尤其重要(A.Tarke等.bioRxiv的预印本https://doi.org/gh6tkp)。 研究小组从志愿者那里收集了T细胞,这些志愿者已经从祖先SARS-CoV-2株感染中恢复过来,或者已经接受了mRNA冠状病毒疫苗。然后,研究人员测试了细胞识别四种新兴变体中蛋白质片段的能力,其中包括最早在南非发现的B.1.351变体。 大部分志愿者的T细胞都能识别所有四个变异体,这要归功于不受变异体突变影响的病毒蛋白片段。结果表明,T细胞可以靶向这些变体。 被COVID-19杀害的人的坟墓在巴西马瑙斯的一座公墓中围着一个工人。图片来源:Michael Dantas /法新社/盖蒂 3月4日-一种新的病毒变体袭击了遭到COVID破坏的城市 在巴西马瑙斯市检测到的冠状病毒变异可能正在推动再感染,并且是该市第二波COVID-19流行。 在大流行的第一波期间,马瑙斯经历了世界上最高的感染率之一:到2020年10月,估计有三分之二的居民被感染,导致一些研究人员预测整个人群的免疫力可能会导致新的感染减少。但是在2021年1月,研究人员在该市住院率上升期间发现了一种新型冠状病毒变种,称为P.1,并将该变种与一些再感染病例相关联。 为了进一步表征该变体,伦敦帝国学院的Nuno Faria和他的同事分析了11月至12月之间从Manaus的184个人类样本中收集的病毒基因组(NR Faria等人的预印本,网址为https://go.nature.com/3sor3jj ; 2021)。该变体包含17个可改变SARS-CoV-2蛋白的突变。在这些变化中,SARS-CoV-2刺突蛋白的变化以前与传播增加和免疫逃逸有关。 通过对P.1的扩散及其在马瑙斯第二波中可能产生的影响进行建模,研究人员估计该变体的传播能力是其他谱系的1.4-2.2倍,并且能够逃避先前感染所赋予的某些免疫力。调查结果尚未经过同行评审。 点击查看:更多有关冠状病毒文章更多医学分类文章使用文档翻译功能 免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:nature
2021-03-11 20:42:46

加强的测序工作正在帮助识别可能促进传播或帮助病毒逃避免疫反应的突变。
埃文·卡拉威(Ewen Callaway)
驾车者在美国最大的COVID测试站点排队,位于加利福尼亚洛杉矶的道奇体育场外。信用:赵林歌/祖玛线
对于过去一年研究了数十万个冠状病毒基因组的科学家而言,美国一直是一个谜。尽管拥有世界领先的基因组测序基础设施,并且比其他任何国家都遭受更多的COVID感染,但美国直到最近才在冠状病毒基因组测序和发现令人担忧的变体方面远远落后。
但是最近几周,美国研究人员发现了许多新的变种,包括在加利福尼亚,纽约州,路易斯安那州和其他地方。他们将继续加大SARS-CoV-2测序工作的力度。这就带来了另一个挑战:弄清所发现的变体。它们带有潜在的令人担忧的突变,并且可能会变得越来越普遍,但是缺乏有关变种如何传播的数据意味着它们所构成的威胁尚不清楚。
资料来源:CDC
“这是狂野的西部,”位于什里夫波特的路易斯安那州立大学健康科学中心的病毒学家杰里米·卡米尔(Jeremy Kamil)说,他领导了一个团队,该团队上个月在路易斯安那州,新墨西哥州和其他地方发现了一种快速上升的变种。卡米尔说,在缺乏有关变体行为的清晰数据的情况下,“似乎有一种非正式的政策,即每个变体都是值得关注的变体,除非另行证明”。
挑战的一部分是美国冠状病毒测序和监测工作的分散性。纽约市哥伦比亚大学的病毒学家戴维·霍(David Ho)说:“现在是各个实验室,州或城市发挥作用”,他的团队上周在该市发现了一个变异体,该变异体可能破坏免疫反应。由于这项零星的工作,纽约,加利福尼亚和华盛顿等州分别贡献了数千个序列,而爱荷华州,田纳西州和新罕布什尔州等其他州则从少得多的COVID病例中获得了序列。
资料来源:CDC
何和其他美国研究人员羡慕英国,该国与公共卫生,医学和研究机构紧密合作以识别变异的全国测序工作已经产生了300,000多个冠状病毒基因组。由于其数据的精细性,英国在2020年底的研究表明,名为B.1.1.7的变体的传播速度明显快于其将要取代的循环菌株。随后的研究表明,B.1.1.7可能更致命,但不会危害疫苗。
是否想更快地追踪流行病变种?解决生物信息学瓶颈
何说:“我认为我们没有那样的东西。” 他希望,美国国立卫生研究院和疾病预防控制中心(CDC),负责生物医学研究和公共卫生的联邦机构,“将使该国更加协调一致地前进”。由疾病预防控制中心(CDC)领导的一项工作于11月启动,目标是每周对大约7,000个样本进行测序(该目标在2月下旬首次实现),最终达到25,000。
令人担忧的突变
在缺乏明确的流行病学或医学数据的情况下,科学家们可以通过携带的变异来评估变异的某些潜在威胁。根据实验室和流行病学研究,研究人员已经提出了越来越多的突变列表,这些突变可能会增强传播或帮助病毒逃避免疫反应。
Ho的团队在纽约发现的变种(也称为B.1.526)带有一个臭名昭著的突变E484K,该突变已在南非和巴西发现的变种中发现。多个实验室的研究表明,E484K的变化(位于识别宿主细胞的冠状病毒刺突蛋白的一部分中)削弱了通常可以使病毒失活的抗体的效力。这可以帮助解释这样的观察结果:在南非和巴西,类似的变体落后于再感染病例,并且在田间试验中降低了疫苗效力。
基于这些担忧,由Ho和哥伦比亚大学微生物学家Anne-Catrin Uhlemann领导的团队建立了一个监视网络,以识别纽约市携带E484K的病毒。B.1.526变体的第一批病例出现在11月,到1月中旬增长到该市总病例的5%,到2月增长到12%。在公共测序数据库中,研究人员发现了B.1.526在美国东北海岸的上下游,甚至远至新加坡。E484K的声名狼藉还激发了帕萨迪纳加州理工学院结构生物学家Pamela Bjorkman和Anthony West带领的团队研究公共测序数据,发现了在纽约出现的血统。
Ho承认B.1.526需要更多的研究。尚未显示出它可以躲避免疫反应,并且其频率的明显升高可能与任何生物学特性无关。“英国变种[B.1.1.7]花费了几个月的时间才显示出更具传播性和更强的毒性。我认为我们也需要这样做。”他说。
罕见的变体
加强美国测序工作正在寻找具有新的或罕见的突变的变体,这些变体更难理解。卡米尔(Kamil)与新墨西哥州的研究人员合作,是因为他们还观察到了由一个被称为鹈鹕的变异引起的病例数上升,这种变异带有从未见过的突变。他们在美国测序数据中发现了其他类似变化的变体。(所有人都被赋予了小鸟的昵称,包括罗宾,Yellowhammer和知更鸟。)
名为Q677P的突变位于刺突蛋白的一个区域附近,该区域需要卡扣成两个,以使病毒颗粒进入宿主细胞。该区域的突变发生在快速传播的变种中,例如B.1.1.7,但卡米尔说,鹈鹕变种目前是值得关注而不是担心的变种。他说:“现在还没有科学的信心说这是一个特别令人担忧的突变,还为时过早。”
上周,加利福尼亚的研究人员对那里发现的带有刺突蛋白突变(称为L452R 1)的变体提出了警告。加利福尼亚大学旧金山分校(UCSF)的一个研究小组发现,具有该突变的变异体在一个城市附近迅速上升,从11月的16%测序样本中出现到1月中旬的一半以上。据媒体报道,另一个加州大学旧金山分校的团队在实验室测试中发现,具有L452R突变的变体更具感染性,对抗体的敏感性更低。
但是许多研究人员对L452R变体的重要性表示怀疑。惠康统计统计学家杰弗里·巴雷特(Jeffrey Barrett)说,这种突变尚未在实验室研究中出现,该研究已标记出其他令人担忧的变化,例如E484K,并且相同的L452R突变已在美国其他地方突然出现,并且增长并不迅速。英国欣克斯顿的桑格研究所。他说:“从根本上讲,这些问题可能不会成为其中之一。” “这是观望加利福尼亚和其他地方情况的问题。”
虽然研究人员试图弄清新近发现的美国变体,但加倍的测序工作也使更多与全球关注的变体有关的病例增多。到目前为止,美国的研究人员仅发现了与南非和巴西发现的逃避免疫变异有关的少数病例。但是在英国发现的B.1.1.7变种的案例正在稳步上升-在欧洲和中东的其他国家中也屡见不鲜。
研究人员说,哪种变体将占主导地位,研究人员说,但是随着疫苗接种的增加,诸如B.1.1.7之类的易感变体可能会减少,而能够部分逃避免疫力的那些变体可能会引发区域性爆发。巴雷特说:“我认为我们不会有多年的'纽约变体'和'加利福尼亚变体'。弄清楚正在发生的事情,不仅取决于对更多样本进行测序,还取决于建立理解样本的能力。何说:“美国必须在这些方面做得更好。”
点击查看:更多有关冠状病毒文章
更多医学分类文章
使用文档翻译功能
免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。
来源于:nature
2021-03-08 14:25:23

Thapsigargin是主要人类呼吸道病毒的广谱抑制剂:冠状病毒,呼吸道合胞病毒和A型流感病毒 经过: 莎拉·贝塔吉(Sarah Al-Beltagi)克里斯蒂安·亚历山德鲁·普雷达利亚·古尔丁乔·詹姆斯Pu娟保罗·斯金纳江志敏王琳琳杨佳云阿什利C.肯尼斯·梅利兹帕维尔·格什科维奇(Pavel Gershkovich) 6,克里斯托弗·J·海斯乔纳森·阮·范·塔姆伊恩·布朗刘金华和张建洲1.诺丁汉大学动物医学与科学学院,萨顿·博宁顿,诺丁汉LE12 5RD,英国2.英国沃金GU24 0NF,Pirbright,Ash Road,Pirbright学院3.英国Addlestone KT15 3NB,Woodham Lane的Weybridge,动植物卫生局(APHA)4.中国农业大学动物医学院,农业部动物流行病学重点实验室,北京圆明园西路2号,北京1001935.英国诺丁汉萨顿博宁顿诺丁汉大学生物科学学院,LE12 5RD,英国6.英国诺丁汉大学公园诺丁汉大学药学院NG7 2RD,英国7.英国诺丁汉大学公园诺丁汉大学化学学院NG7 2RD,英国8.英国诺丁汉大学公园诺丁汉大学医学院NG7 2RD*应与之联系的作者。这些作者为这项工作做出了同等的贡献。 摘要:SARS-CoV-2和其他主要呼吸道病毒的长期控制策略除了明智使用有效疫苗外,还需要包括抗病毒药以治疗急性感染。虽然正在为大规模疫苗接种推出COVID-19疫苗,但针对任何疾病使用或开发的适量抗病毒药物证明了抗病毒开发的挑战。我们最近发现,无毒的毒胡萝卜素(TG)是肌腱蛋白/内质网(ER)Ca2 + ATPase泵的抑制剂,可诱导有效的宿主先天免疫抗病毒反应,从而阻止甲型流感病毒的复制。在这里,我们显示TG还可以在永生或原代人细胞中阻断呼吸道合胞病毒(RSV),普通感冒冠状病毒OC43,SARS-CoV-2和A型流感病毒的复制。 TG的抗病毒性能在抑制OC43和RSV方面显着优于瑞贝昔韦和利巴韦林。值得注意的是,TG在合并感染中对冠状病毒(OC43和SARS-CoV-2)和流感病毒(苏联H1N1和pdm 2009 H1N1)的抑制作用相同。酸稳定TG的感染后口服管饲法可保护小鼠免于致命的流感病毒攻击。结合其在主动感染之前或期间抑制不同病毒的能力,以及在TG暴露后至少48小时的抗病毒持续时间,我们建议TG(或其衍生物)是有希望的广谱抗SARS-CoV抑制剂-2,OC43,RSV和流感病毒。 关键字:毒胡萝卜素;抑制剂抗病毒物质; SARS-CoV-2;冠状病毒OC43;呼吸道合胞病毒;流感病毒广谱先天免疫;老鼠;瑞地昔韦利巴韦林;奥司他韦 1. 介绍在COVID-19大流行中,针对大规模疫苗接种的有效疫苗的需求从未如此大,以控制和预防可导致医院不堪重负的猖ramp的发病率,死亡率和激增的临床病例。 COVID-19是由一种新型冠状病毒引起的,该冠状病毒被称为严重急性呼吸系统综合症冠状病毒2(SARS-CoV-2),一种包膜的阳性单链RNA病毒[1]。尽管现在正在部署COVID-19疫苗,但根除是不切实际的,并且对有效抑制SARS-CoV-2抗病毒药[2,3]的平行需求存在着并行的需求,以抑制患者体内主动病毒复制以逆转病毒进程。考虑到并不是所有的疫苗都可以在随后的感染中预防病毒的脱落,因此抗病毒药也可以用来降低人群中传播的总体病毒水平,从而减少其传播。针对任何传染病使用或开发的抗病毒药物数量很少,证明了抗病毒药物开发的实际挑战。特别是对于呼吸道病毒,通常遇到的主要障碍是病毒突变抗性的出现,这种突变抗性源于直接针对病毒特定部分的普遍采用的策略。甲型流感病毒的N1部分对病毒神经氨酸酶抑制剂奥司他韦的耐药性日益引起人们的关注[4,5]。人们对令人担忧的是对新近引入的针对病毒PA蛋白的流感抗病毒药baloxavir marboxil的耐药性不断提高[6];甲型流感病毒的PB1基因中的突变很容易赋予对新型核苷类似物抗病毒药物favipiravir的耐药性[7]。可以潜在地规避病毒突变挑战的另一种抗病毒设计方法是,在早期感染期间调节通用的宿主固有免疫反应,以充分破坏病毒复制,从而建立获得性免疫(抗体和细胞介导的)。这种新颖的以宿主为中心的抗病毒方法可以具有抑制不同类型病毒的额外好处[8]。鉴于表现出不同病因的急性呼吸道病毒感染在临床表现上是无法区分的,因此可以同时针对不同病毒类型的广谱以宿主为中心的抗病毒药可能会大大改变临床管理。除了目前的SARS-CoV-2大流行外,甲型流感病毒和呼吸道合胞病毒(RSV)仍然是造成人类呼吸道病毒感染的重要因素。流感病毒是包膜的负义分段单链RNA病毒,每年在全球范围内导致多达65万例死亡,并且是造成先前大流行的原因[9]。 RSV是一种包膜的负义单链RNA病毒,是幼儿和老年人急性下呼吸道感染的主要原因,与5岁以下儿童的院内死亡48,000–74,500相关,其中99%在发展中国家发生的死亡[10,11]。虽然在甲型流感病毒感染中使用抗病毒药可能会受到病毒抗药性的阻碍,但仅批准使用抗RSV的化学抗病毒药利巴韦林仅限于婴儿的严重感染[5,12]。我们最近显示,毒胡萝卜素(TG)是肌浆网/内质网(ER)Ca2 + ATPase泵的抑制剂[13],在纳摩尔非细胞毒性水平上诱导了有效的宿主抗病毒应答,从而阻断了甲型流感病毒的复制[14]。TG诱导的ER应激未折叠蛋白反应(UPR)似乎是激活宿主抗病毒防御下游谱的主要驱动力。在这里,我们显示TG实际上是一种以宿主为中心的广谱抑制剂,在永生或原代人类细胞中,可有效阻断RSV,普通感冒冠状病毒OC43,SARS-CoV-2和A型流感病毒的复制。我们还确定,胃中发现的TG是低pH稳定的,从一次引发起就能够快速诱导持续至少48小时的有效抗病毒状态,并且在小鼠口服后可在小鼠体内进行治疗性保护(体内)感染,应对致命的流感病毒挑战。 2. 材料和方法2.1. 细胞和病毒原代正常人支气管上皮细胞(NHBE)和支气管上皮生长培养基由Promocell(德国海德堡)提供。在添加了10%胎牛血清(FCS)的DMEM-Glutamax中培养无限增殖的新生猪气管上皮(NPTr)细胞[15],HEp2细胞,MRC5细胞,A549细胞,Calu-3细胞,Vero E6细胞和MDCK细胞。100 U / mL青霉素-链霉素(P/S)(Gibco,ThermoFisher Scientific,佩斯利,英国)。通过DEFRA分离出的人RSV(A2株,ATCC VR-1540),苏联H1N1(A / USSR / 77),PR8 / H1N1(A / PR8 / 1934)和pdm H1N1病毒(A / Swine / England / 1353/2009)这项研究使用了SwIV监视程序SV3401)。还使用了人冠状病毒OC43(OC43)和SARS-CoV-2病毒(2019-nCoV /意大利-INMI1株,从EVAg获得的进化枝V(008V-03893)),后者的完整序列已提交给GenBank(SARS-CoV- 2 / INMI1-Isolate / 2020 /意大利:MT066156),并且可以在GISAID网站上找到(betaCoV /意大利/ INMI1-isl / 2020:EPI_ISL_410545)。 2.2. 细胞活力测定根据制造商的说明,使用CellTiter-Glo发光细胞活力测定试剂盒或CellTiter-Glo 2.0细胞活力测定试剂盒(Promega,麦迪逊,威斯康星州,美国)确定细胞活力。 2.3. 细胞化学启动根据制造商的建议,将TG(美国密苏里州圣路易斯的默克公司)和瑞德昔韦(德国慕尼黑的Selleckchem公司)溶解在DMSO中,而利巴韦林和羟氯喹(默克公司)按照制造商的建议溶解在水中。化合物的抗病毒方案对细胞活力没有可检测到的不利影响。通常,除非另有说明,否则在感染前将细胞在适当的细胞培养基中稀释的所示化合物中孵育(稀释30分钟内使用),然后孵育30分钟,然后用PBS洗涤3次并用所示病毒感染(感染前启动)。或者,首先将细胞感染指定的时间,然后将化合物灌注30分钟,然后继续进行感染培养(感染后灌注)。细胞的所有TG引发仅持续30分钟,然后根据特定实验用PBS洗涤并进行下游培养。2.4. RSV感染和子代病毒定量HEp-2细胞,A549细胞和NHBE细胞根据24小时噬斑测定,在补充2%FCS的DMEM-Glutamax中以指定的RSV感染复数(MOI)感染48或72小时,收集的离心上清液用于噬菌斑测定或RT-qPCR的病毒拷贝数进行定量(请参见2.6节)。以前已经描述了通过噬斑分析对HEp2细胞进行RSV滴定[16]。简而言之,将感染细胞上清液的连续稀释液(一式三份)连续感染24小时的HEp2细胞固定在丙酮:甲醇(1:1)中,并使用小鼠抗RSV(2F7)抗体(1:1000)进行免疫染色(Abcam (英国剑桥)。2.5. 流感和冠状病毒的单一和共感染,以及子代病毒的定量甲型流感病毒感染需要无血清培养基和胰蛋白酶。 NPTr细胞和A549细胞的感染培养基是Opti-MEM I(Gibco),辅以100 U / mL P / S,2 mM谷氨酰胺和200 ng / mLL-1-甲苯磺酰胺-2-苯乙基氯甲基酮( TPCK)胰蛋白酶(Sigma-Aldrich)。根据病灶形成试验(FFA),将特定的流感病毒MOI在感染培养基中感染细胞2到3 h,然后用PBS洗涤3次,并在新鲜感染培养基中孵育24天如图所示,持续72小时。收集纺丝上清液用于通过6 hFFA,50%组织培养物感染剂量(TCID50)或通过RT-qPCR的病毒拷贝数对子代病毒进行定量。以前已经描述了通过FFA滴定流感病毒[14]。在添加100 U/ mLP / S,2 mM谷氨酰胺和100 ng / mL TPCK胰蛋白酶的Opti-MEM I中,将A549细胞和MRC5细胞用指定的MOIOC43(基于FFA)感染OC43 3小时。用PBS洗涤3次,并在新鲜的感染培养基中孵育长达72小时。对用OC43和苏联H1N1病毒共感染的A549细胞进行了相似的处理。收集纺丝上清液用于通过FFA定量子代病毒或通过RT-qPCR定量病毒拷贝数。 在指示的MOI上,以TCID50为基础,用SARS-CoV-2将Calu-3,NHBE和Vero E6细胞感染,在补充有100U/mLP/ S,2 mM谷氨酰胺和100ng的Opti-MEMI中感染3 h。如图所示,将1mL / mL TPCK胰蛋白酶,随后在PBS中洗涤3次,并在新鲜的感染培养基中孵育总计长达72小时。对被pdm H1N1病毒和SARS-CoV-2病毒共感染的Calu-3细胞进行了相似的处理。收集纺丝上清液用于通过TCID50定量子代病毒或通过RT-qPCR定量病毒拷贝数。以96孔板形式进行TCID50滴定,以定量SARS-CoV-2和A型流感病毒,最少重复四次。在生长培养基DMEM(Gibco)中对每个样品进行十倍稀释。然后,将50µL稀释样品分别添加到MDCK和VeroE6细胞中用于流感和SARS-CoV-2滴定,并在37°C下孵育1 h。然后将培养基换成200 µL DMEM(补充200ng /mL TPCK胰蛋白酶用于流感滴定),并在37℃下孵育5天。在显微镜下评估每个孔的细胞病变作用。使用Spearman-Karber方法以TCID50计算病毒滴度[17,18]。检测限为5.62 TCID50 / mL。2.6. RNA制备和实时RT-PCRRNeasy Plus Minikit(Qiagen,希尔登,德国)用于从细胞中提取总RNA。使用Superscript III First Strand合成试剂盒(ThermoFisher Scientific)从1 µg总RNA合成cDNA。用LightCycler 96仪器(Roche,Basel,Switzerland)进行宿主基因的表达。基因表达的计算基于比较的Ct方法,已标准化为18S核糖体RNA。 HSPA5(FH1_HSPA5和RH1_HSPA5),HSP90B1(FH1_HSP90B1和RH1_HSP90B1)和DDIT3(FH1_DDIT3和RH1-DDIT3)的人ER应力引物;和人类IFNB1引物(FH1_IFNB1和RH1_IFNB1),OAS1引物(FH1_OAS1和RH1_OAS1)和RNASEL引物(FH1_RNASEL1和RH1_RNASEL1)是预先设计的Sigma-Aldrich。使用PrimerExpress 3.0.1(ThermoFisher Scientific)设计了其他引物(由Sigma-Aldrich合成),并在表1中显示。QIAamp病毒RNA试剂盒(Qiagen)用于从离心细胞培养上清液中提取病毒RNA。使用QIAGEN OneStepRT-PCR KIT进行一步实时qPCR,以定量相对病毒拷贝数。 表1.引物序列。 基因没有第一(5d – 3d)反义引物(5j–3j)18S核糖体RNA(通用)ACGGCTACCACATCCAAGGACCAATTACAGGGCCTCG-AAA F基因(RSV)CAAGAACTGACAGAGGATGGTACTGCATGTTTCAGCTTGTGGGAAGAL基因(RSV)AACACTTATCCTTCTTTGTTGGAACTTAGCAACCGAAACTCACGATAGAAAM基因(RSV)ACTCAAGAAGTGCAGTGCTAGCAAAGGACACATTAGCGCATATGGTRIG-I(人类)GAAGGCATTGACATTGCACAGTTGGTTTGGATCATTTTGATGACAM基因(苏联H1N1)AGATGAGCCTTCTAACCGAGGTCGTGCAAAAACATCTTCAA-GTCTCTGM基因(pdm H1N1)AGATGAGTCTTCTAACCGAGGTCGTGCAAAGACACTTTCCA-GTCTCTGOrf1ab(SARS-CoV-2)CCGATCATCAGCACATCTAGGTTGACAAGGCTCTCCATCT-TACCTTTOrf1ab(OC43)GCCAGGGACGTGTTGTATCCTTGATCTTCGACATTGTGACCTATG 2.7. 蛋白质印迹使用放射免疫沉淀测定(RIPA)缓冲液(圣克鲁斯,达拉斯,美国德克萨斯州)补充1%苯基甲基磺酰氟(PMSF)(圣克鲁斯),1%抑制剂混合物和1%原钒酸钠,裂解细胞,并通过Bio-RAD蛋白质测定法(Bio-Rad,Hercules,CA,USA)测量蛋白质浓度。一抗是1:500稀释的山羊抗甲型流感A1(Abcam,ab20910),小鼠抗甲型M2克隆14C2(1:1000)(Invitrogen,MA1082),山羊抗甲型流感病毒多克隆抗体以1:2000的稀释度(Abcam,ab155877),小鼠抗冠状病毒抗体OC-43株,克隆541–8F处于1:1000(Sigma-Aldrich,MAB9012)和小鼠抗β-肌动蛋白克隆AC- 74处于1:5000(Sigma-Aldrich,A2228)。合适的物种特异性二抗是过氧化物酶偶联的(Abcam),用于化学发光检测(Amersham ECL Western Blotting Detection Reagent,美国马萨诸塞州马尔伯勒)。2.8. 干扰素-β(IFNβ)ELISA通过酶联免疫吸附测定(ELISA)定量细胞培养上清液中的IFNβ分泌。 IFNβELISA(人类IFN-βQuantikine ELISA试剂盒,美国明尼苏达州明尼阿波利斯市Bio-Techne)按照制造商的说明进行。样品进行三次重复分析。2.9. 小鼠流感病毒挑战为了评估TG在感染过程中(即感染后)的抗病毒效力,并与体内奥司他韦进行比较,将6至8周龄的BALB / c小鼠(雌性)分为两组,以确定感染后存活率(每个治疗组n = 8)和子代病毒产生(每个治疗组n = 8)。小鼠经鼻内感染了3个MLD50的PR8/H1N1病毒。感染后十二小时,每天通过管饲法口服TG(1.5 µg / kg /天),奥司他韦(45 mg / kg /天)或PBS + DMSO(PBS-DMSO对照中DMSO的百分率与其他治疗相当)天。在感染后第3和5天,收集每组四只小鼠的肺以进行病毒滴定。通过TCID50分析,对接种了10倍连续稀释的均质化肺组织的MDCK细胞进行病毒滴定,并在37°C下孵育72小时。 TCID50值是根据Reed–Muench方法[19]计算的。 2.10. 量化与统计分析使用GraphPad Prism 7和图例中描述的统计方法进行统计分析。 p值<0.05被认为是显着的,并表示为* p <0.05,** p <0.01,*** p <0.001和**** p <0.0001。 Kaplan–Meier方法用于生存分析。给出的结果代表了三个或更多个独立的重复,除非另有说明,否则误差线为标准偏差。2.11. 道德声明所有动物工作均已获得北京科学技术协会(ID:SYXK(Beijing)2007–0023)的批准,并按照北京实验动物管理委员会发布的《北京实验动物福利和道德规范》进行;符合中国农业大学机构动物护理和使用委员会指南(ID:SKLAB-B-2010–003)。3. 结果3.1. TG阻止子代RSV生产为了证明TG对RSV的抗病毒活性,在感染前24小时(图1A)或感染后24小时(hpi)(图1B)用TG短暂灌注人HEp2和A549细胞30分钟。每种细胞类型的感染前和感染后TG引发导致后代病毒产量显着下降(统计上显着)。感染前用0.5 µM TG引发的HEp2细胞将子代病毒的产生减少了近10,000倍(图1A);在24 hpi时用0.5 µM TG引发的细胞将病毒输出降低约1000倍(图1B),这分别表明其对RSV的预防和治疗抗病毒潜力。用TG对HEp2和A549细胞进行30分钟的单次启动诱导了有效的抗病毒状态,这种状态持续至少48小时(图1C,D)。 TG引发HEp2细胞(感染前即刻或感染前48小时)引发了病毒L,F和M基因的转录抑制(图1E,F)。所用TG的抗病毒方案对HEp2和A549细胞无细胞毒性(图1G,H)。补充证据TG的非细胞毒性抗病毒剂量方案已在我们较早的出版物中发现[14]。因此,TG作为一种非细胞毒性抑制剂具有快速作用,可在持续至少48小时的细胞中诱导抗病毒状态,并且在主动RSV感染之前或期间使用时是有效的。 接下来,根据被感染的HEp2细胞的后代病毒产量(图2A)和病毒RNA检测(图2B),将TG的抗病毒性能与病毒唑进行比较,利巴韦林是一种被批准用于严重RSV感染的幼儿的抗病毒药物。 TG在阻止RSV复制方面优于病毒唑。例如,用0.1µM TG感染HEp2细胞预感染30分钟对子代病毒产生的抑制作用比连续使用30 µM利巴韦林(50%利巴韦林有效浓度(EC50)对RSV = 11)高160倍。 µM [20])(图2A)。 TG在HEp2细胞的RSV抑制中表现出984的选择性指数(50%(CC50)/ EC50的细胞毒性浓度)和84.55 nM的EC90,这表明它具有很强的安全裕度(图2C,D)。在由TG引发和感染的原代NHBE细胞中,相应培养基中病毒RNA的检测降低(图2E)与病毒L,F和M基因的转录抑制(图2F)同时发生,这进一步表现为TG剂量依赖性减少病毒蛋白的产生(图2G,H)。 两者合计,TG作为抗病毒剂比利巴韦林更有效,表现出很强的选择性指数,并阻断了RSV病毒的转录和病毒蛋白的产生。在原代NHBE细胞中,相对于DMSO对照,在RSV感染之前和期间,TG依赖于TG的剂量以TG剂量依赖性的方式进行了TG的感染前致敏增强ER应激基因(DDIT3,HSPA5和HSP90B1)的表达(图3A,C)。TG还增加了NHBE细胞中RIG-I信号相关基因(RIG-I,IFNB和RNASEL)的基础表达,但是在感染过程中,相对于被感染的DMSO对照,RIG-I相关基因的诱导显着衰减(图3D,G)。因此,在NHBE细胞中RSV感染期间,RIG-I相关基因的转录诱导减少是一个特征。TG介导的RSV抑制作用。3.2. TG阻止子代冠状病毒OC43的生产为了证明TG对冠状病毒的抑制作用,在感染地方性OC43病毒之前,立即用TG引发A549细胞30分钟[21](图4A–C)。以剂量依赖性方式,TG阻断了病毒复制,如感染细胞的培养基中病毒RNA的急剧减少(图4A),病毒转录的抑制(图4B)和病毒NP蛋白生成减少(图4C)所证明。将TG的抗病毒性能与羟氯喹(HC)进行了比较,后者是一种毒性比氯喹低的化合物,可抑制OC43[22]。 TG在阻止子代OC43病毒产生方面比HC更有效(图4D,E)。感染前引发与整个感染过程中连续使用20 µM HC相比,0.05 µM TG仅持续30分钟对OC43的抑制作用更大(针对SARS-CoV-2的HC EC50 = 4.51 µM [23])(图4D)。与HC处理后代病毒的适度减少不同,在TG引发的感染细胞的培养基中几乎未检测到任何病毒(图4E)。在A549细胞中,TG在阻止OC43病毒(图4F)和流感病毒(图4G)复制方面比最近被批准用于SARS-CoV-2感染紧急使用的核苷类似物Remdesivir(RDV)[24]更好。 。尽管在72hpi时,细胞连续暴露于0.3 µM RDV(RDV EC50对OC43 = 0.15 µM [24])显示约17,000倍减少子代病毒RNA的检测(相对于相应的DMSO对照);细胞反过来,用0.3 µM TG引发的细胞则比RDV处理过的细胞具有450倍的抑制作用(图4F)。在72 hpi时,与0.3μM的RDV连续使用相比,0.05μMTG引发的细胞对苏联H1N1病毒的抑制作用还强于7倍。 RDV对流感病毒复制几乎没有抗病毒作用(图4G)。 TG在A549细胞中的抗病毒用途无细胞毒性(图4H)。 TG在MRC5细胞中OC43上的TG的选择性指数(CC50 / EC50)高,介于7072和9227之间(图4I)。总的来说,TG强烈抑制OC43病毒的转录和蛋白质产生,作为抗病毒剂比HC和RDV更有效,并且具有较高的选择性指数。 点击:查看文章剩下部分 查看更多医学文章 查看更多冠状病毒文章 使用专业文章翻译功能免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。 来源于:mdpi
2021-02-24 16:25:36

Thapsigargin是主要人类呼吸道病毒的广谱抑制剂:冠状病毒,呼吸道合胞病毒和A型流感病毒
点击:查看上部分内容介绍
图1. thapsigargin(TG)在无细胞毒性水平下短时间(30分钟)暴露于人细胞,迅速引起延长的(≥48h)抗病毒状态,从而阻止了RSV复制。 TG在(A)感染前24小时或(B)感染后24小时以剂量依赖性方式引发,可阻止RSV产生。 (A)用TG或对照DMSO灌注HEp2和A549 30分钟,用PBS洗涤并在RSV感染前在正常培养基中孵育24小时,或(B)首先在TG 30分钟之前用RSV感染24小时启动。 TG诱导的抗病毒状态持续至少48小时。如图所示,用TG或DMSO对照将HEp2(C)和A549(D)细胞灌注30分钟,用PBS洗涤,再进行24或48小时的正常培养;之后,细胞被RSV感染。所有细胞均以0.1 MOI感染RSV(A2株,ATCC VR-1540),共3天。收集离心的上清液感染HEp2细胞24小时,用小鼠抗RSV(2F7)抗体(pfu / mL)免疫检测RSV。2向ANOVA(Sidak的多次比较)的意义与相应的DMSO控件有关。与A549细胞相比,HEp2细胞更易于RSV复制。 TG抑制RSV转录。在RSV感染之前,立即用0.5µMTG灌注HEp2细胞(E)30分钟,在RSV感染之前(F)用TG灌注HEp2细胞30分钟,用PBS洗涤并培养48小时。总共感染3天后,提取总RNA进行cDNA转换,以量化标准化为18srRNA的病毒基因(L基因,M基因和F基因)表达。通过相对于相应DMSO对照中表达的未配对t检验确定显着性。 (G)用指定浓度的TG或DMSO对照预孵育30分钟的(Hp2和(H)A549细胞)用PBS洗涤,在无血清培养基(Opti-MEM)中培养过夜,并进行细胞活力测定(CellTiter-Glo®发光细胞活力测定试剂盒,Promega)。水平条=平均值(红色)±标准偏差;ns =不重要。与Dunnett进行多次比较的单向方差分析的意义是相对于相应的DMSO控件而言的。所有测定均一式三份,进行三次。 *p <0.05和**** p <0.0001。
图2. TG作为抗病毒药比利巴韦林更有效,显示出高选择性指数并阻止病毒转录和病毒蛋白产生。如前所述,在培养基中或在单独培养基中连续存在利巴韦林时,将HEp2细胞用TG,利巴韦林或DMSO对照引发30分钟,如前所述用PBS洗涤并用RSV感染(用于TG或DMSO对照)。以4 dpi收获培养基,用于(A)子代病毒滴定(pfu /mL)和(B)通过一步反转录-qPCR检测病毒L基因RNA。除非另有说明,否则单向ANOVA(Tukey的多次比较)的显着性是相对于DMSO控件而言的。在HEp2细胞的RSV感染中,TG的相应选择性指数(SI)估计为984(C,D)。在TG的感染前引发范围内,分别通过pfu / mL病毒滴定和发光细胞活力测定法确定HEp2细胞中TG对RSv的有效浓度(EC)和细胞毒性浓度(CC)。 SI = CC50 / EC50 = 63.43 / 0.06447 = 983.9。 EC90 = 84.55 nM。如图所示,用TG或DMSO对照引发NHBE细胞30分钟,用PBS洗涤并以0.1 MOI感染RSV。在48 hpi时,通过一步反转录qPCR(E)收集用于检测病毒L基因RNA的培养基,提取总RNA进行cDNA转化以定量病毒基因的表达(L基因,F -基因和M基因)标准化为18s rRNA(F)。单向ANOVA(Dunnett的多重比较)(E)和两向ANOVA(Tukey的多重比较)(F)的意义与DMSO控件有关。 TG还抑制NHBE细胞中病毒F蛋白的产生。如图所示,用TG或DMSO对照灌注细胞30分钟,用PBS洗涤,并在80℃下用RSV感染
0.05 MOI持续48小时,并直接免疫染色是否存在RSV F蛋白(G,H)。以100倍放大率拍摄的图像。随着TG启动剂量的增加,RSV阳性细胞(pfu)的数量明显减少。单向方差分析(Dunnett的多次比较)的意义与DMSO控件有关。 * p <0.05,*** p <0.001和**** p <0.0001。
图3. NHBE细胞的TG引发增加了ER应激基因的表达(感染前和感染后)和RIG-I信号传导相关基因的基础表达,但是在感染过程中,RIG-I关联基因的诱导是衰减。用TG或DMSO对照引发细胞30分钟,用PBS洗涤并在0.05 MOI下用RSV感染48小时,然后提取总RNA进行cDNA转化以定量ER应激基因的表达(A)DDIT3,(B)HSPA5 (C)HSP90B1。将表达标准化为18s rRNA,通过2向ANOVA(Sidak的多次比较)确定的显着性相对于相应的DMSO对照。有一致的迹象表明,TG启动引发了RIG-I相关基因RIG-I(D),IFNB(E)(以及相应的IFNB蛋白(F))和RNASEL(G)的感染前激活。在16hpi对上清液进行IFNB ELISA。将所有RNA表达标准化为18s rRNA,并通过单向ANOVA和Dunnett的多重比较(D)或2向ANOVATukey的多重比较(E–G)确定的显着性相对于相应的DMSO对照。所有测定均一式三份,进行三次。*p <0.05,** p <0.01,*** p <0.001和**** p<0.0001。
图4. TG抑制OC43病毒的转录和蛋白质的产生,作为抗病毒剂比羟氯喹和瑞姆昔韦更有效,并且显示出高选择性。 (A,B)用TG引发A549细胞30分钟,用PBS洗涤两次,并在0.5MOI下用OC43感染3小时。然后,将细胞再次用PBS洗涤,并在无血清培养基(补充有0.1 µg / mL TPCK胰蛋白酶的Optii-MEM)中培养48小时,然后从(A)培养基和(B)细胞中提取RNA,分别进行反转录qPCR和cDNA转换,再进行qPCR,以检测病毒OC43复制酶多蛋白1ab RNA。 cDNA定量标准化为18s rRNA。 (C)使用重复的孔组进行细胞蛋白提取,以通过蛋白质印迹法检测OC43 NP。 (D,E)如图所示,用TG,羟氯喹(HC)或DMSO / PBS对照对MRC5细胞进行预处理,持续30分钟,用PBS洗涤两次,用OC43(0.01 MOI)感染3 h,再用PBS两次,最后在无化合物(感染前)或持续存在HC(连续)的情况下用无血清培养基补充。在2 dpi时,收集培养基用于(D)通过一步反转录qPCR检测病毒多蛋白1abRNA和(E)通过感染A549细胞24小时并免疫染色是否存在病毒NP来直接检测子代病毒。以100倍放大率拍摄的图像。所指示的显着性(通过单向方差分析确定)和病毒RNA检测的降低百分比相对于相应的对照。 (F,G)TG在阻止OC43(F)和甲型流感病毒(G方面优于伦德韦(RDV))复制。用指示的TG,0.3 µM RDV或DMSO对照对A549细胞进行初次免疫30分钟,用PBS洗涤两次,并用0.01 MOI的CoVOC43或1.0 MOI的苏联H1N1病毒感染2小时,然后再次用PBS,然后在无血清培养基中孵育TG引发的细胞,或在连续存在RDV的培养基中孵育。在24、48和72 hpi时,基于相对Ct方法,对收集的上清液进行病毒RNA提取,然后进行一步反转录qPCR,以检测OC43复制酶多蛋白1ab RNA或流感M基因RNA的相对拷贝数。 。所指示的显着性是相对于基于2通ANOVA Dunnett(F)或Tukey(G)多重比较测试的RDV处理过的细胞而言的。 (H)RDV和TG处理对细胞活力没有不利影响。将A549细胞用RDV连续处理,或用指定的TG或DMSO处理30分钟,洗涤,培养过夜,然后进行细胞活力测定(CellTiter-Glo 2.0细胞活力测定,Promega)。通过单因素方差分析相对于DMSO对照确定指示的显着性。 (I)TG对OC43的抑制作用的选择性指数(CC50 / EC50)估计在7072和9227之间。EC90= 0.02622 µM。用TG(0至91 µM)灌注MRC5细胞30分钟,用PBS洗涤两次,并在含10%FCS和1%P / S的DMEM Glutamax中培养过夜。用CellTiter-Glo 2.0细胞活力测定(Promega)进行细胞活力测定(CC50)。有效或抑制TG的剂量反应(EC50)是基于以指定浓度的TG(0至0.5 µM)灌注MRC5细胞30分钟,然后进行PBS洗涤和以0.01 MOI的OC43感染。感染后三天,收集上清液用于RNA提取和一步反转录qPCR以定量病毒RNA(多蛋白1ab RNA)的存在。 CC=细胞毒性; EC =有效浓度。 ** p <0.01,*** p<0.001和**** p <0.0001。
接下来,评估了在OC43病毒感染之前和期间对TG启动的ER应激反应。在A549细胞中,TG以剂量依赖性方式基础上和在感染过程中刺激ER应激基因的表达。 TG诱导的A549细胞OC43感染的ER应激基因图谱(图5A,C)与NHBE细胞的RSV感染的TG应激基因图谱高度相似(图3A,C)。在A549细胞中,TG启动似乎对RIG-I相关基因(RIG-I,IFNB和OAS1)的基础转录几乎没有影响。像在NHBE细胞的RSV感染中(图3D,G)一样,在TG引发的细胞的OC43感染过程中,RIG-I相关基因的诱导也减弱了(图5D,F)。因此,在OC43感染期间降低RIG-I相关基因的转录诱导也是TG介导的OC43病毒抑制的特征。重要的是,感染前TG引发的A549细胞与OC43病毒和苏联H1N1甲型流感病毒的共同感染(图5H)一样,能够抑制单独的病毒感染(图5G)。总之,A549细胞的TG引发在基础上和OC43感染期间增加了ER应激基因的表达,在感染期间减弱了RIG-1信号相关基因的诱导,并抑制了与OC43和流感病毒的共感染。
1.1. TG阻止子代SARS-CoV-2生产
SARS-CoV-2与OC43病毒一样易受TG抑制。用TG对Calu-3和NHBE细胞进行感染前预感染可阻断SARS-CoV-2复制(图6A,C),这与在A549和MRC5细胞中用OC43病毒所见的抑制作用相当(图4A,D)。在Calu-3细胞中,用SARS-CoV-2在TG感染24小时后用TG引发感染后30分钟也有效抑制病毒,表明其在SARS-CoV-2感染中具有治疗潜力(图6B)。与流感病毒抑制一样[14],TG无法抑制SARS-CoV-2在Vero E6细胞中的复制,这表明完整的I型IFN系统对于TG介导的宿主抗病毒应答是必需的(图6D)。
通过子代病毒RNA检测(图6E–H)和传染性子代确定,TG的广谱抗病毒效力在单独的病毒感染中与在SARS-CoV-2和pdm H1N1病毒共感染中一样明显在感染的Calu-3细胞的培养基中通过TCID50分析检测病毒(图6I–K)。在72 hpi下,
在单个病毒中,相对于相应的DMSO对照,0.5 µM TG引发的细胞相对于相应的DMSO对照,抑制SARS-CoV-2后代病毒产生300倍(99.7%)和880倍(99.9%)。
感染(图6I)和共同感染(图6J)。相对于相应的DMSO对照,在以0.5 µM TG引发的共感染细胞72 hpi时,pdmH1N1病毒的产量降低了11倍(93.3%)(图6K)。总体而言,在单病毒比较(图6E,F)和共感染(图6G,H)中,SARS-CoV-2与dd H1N1病毒感染相比,TG引发的后代病毒减少比例更高。这表明SARS-CoV -2比dm H1N1病毒对TG抑制更敏感。总之,TG是一种广谱抗病毒药物,主要针对人类主要呼吸道呼吸道合胞病毒,冠状病毒(特别是SARS-CoV-2)和甲型流感病毒,不能区分单病毒感染和复合病毒感染。
图5. A549细胞的TG引发在基础上和在OC43感染期间增加了ER应激基因的表达,在感染期间减弱了RIG-I信号相关基因的诱导,并抑制了与OC43和流感病毒的共感染。 (A–C)。TG引发似乎以剂量依赖的方式刺激内质网应激基因的表达。 (D–F)TG在感染过程中减弱了RIG-I相关基因的诱导。 A549细胞用TG引发30分钟,用PBS洗涤两次,并在0.5 MOI下用OC43感染3小时;此后,再次用PBS洗涤细胞,并在无血清培养基中培养24小时,然后收集细胞裂解液用于RNA提取和cDNA转化,用于(A)DDIT3,(B)HSPA5,(C)HSP90B1,(D )RIG-1,(E)IFNB和(F)OAS1。将所有表达标准化为18s rRNA。意义相对于基于2向ANOVA(Tukey的多次比较)的相应DMSO控件。 (G,H)在单独的病毒感染或共同感染中,TG在A549细胞中抑制了OC43病毒和苏联H1N1病毒的复制。将细胞用TG灌注30分钟,用PBS洗涤两次,并在OC43病毒和OC43病毒和US H1N1病毒感染下
单病毒感染或共同感染3 h分别为0.01和1.5 MOI(基于FFA);之后,将细胞再次用PBS洗涤两次,并在无血清培养基中孵育。在48 hpi收获培养基用于病毒RNA提取,然后进行一步反转录qPCR,以检测OC43复制酶多蛋白1ab RNA和苏联H1N1 M基因RNA的相对拷贝数。所示的显着性基于2次方差分析Tukey的多次比较,并且病毒RNA检测的减少百分比相对于相应的DMSO对照。所有测定均一式三份,进行三次。 * p <0.05,** p <0.01,*** p <0.001和**** p <0.0001。
图6.在单病毒感染和与pdm H1N1病毒共感染中,TG有效地阻止了子代SARS-CoV-2的产生。 (A,C,D)Calu-3和NHBE细胞的感染前TG引发,但不是Vero E6细胞,有效地抑制了子代病毒的输出。按照指示用TG灌注细胞30分钟,用PBS洗涤两次,并在80℃下感染SARS-CoV-2。
0.01MOI 3小时;之后,将细胞再次用PBS洗涤两次,并在无血清培养基中孵育,该培养基中添加了0.2 µg /mL TPCK胰蛋白酶。在72 hpi的培养基上进行病毒RNA提取。 (B)在24 hpi用TG引发阻断了SARS-CoV-2在Calu-3细胞中的复制。首先以0.01 MOI的浓度将SARS-CoV-2细胞感染细胞24小时,
然后用指示的TG灌注30分钟,用PBS洗涤3次并在无血清培养基中孵育。在48和72 hpi的培养基上进行病毒RNA提取。根据相对Ct方法,对以上分离的所有RNA进行一步反转录qPCR,以检测SARS-CoV-2复制酶多蛋白1ab RNA的相对拷贝数。用TG对Calu-3细胞进行预感染引发可抑制(E)SARS-CoV-2和(F)pdm H1N1病毒的单独感染,以及(G,H)两种病毒的共同感染。用TG或DMSO灌注细胞30分钟,用
将相应的病毒以0.01 MOI的浓度持续2 h,用PBS洗涤3次,然后在无血清培养基中孵育。在感染后指定的时间点,对感染细胞的培养基进行采样以进行病毒RNA提取,以进行一步反转录qPCR,以检测SARS-CoV-2复制酶多蛋白1ab RNA(E,G)和流感的相对拷贝数M基因RNA(F,H)。 (IK)在相似感染的培养物上于24、48和72 hpi采集的培养基样本用于检测
通过在Vero细胞中进行TCID50病毒滴定来检测存活的子代病毒(显示为平均值±SEM)。 (一)在单病毒感染中
SARS-CoV-2,TG表现出剂量依赖性病毒抑制作用。在与SARS-CoV-2和pdm H1N1病毒共感染时,TG能够同时抑制SARS-CoV-2(J)和pdm H1N1病毒(K)。所示的显着性基于2向ANOVA Tukey的多次比较测试和相对于相应DMSO对照的病毒RNA变化百分比。 * p <0.05,** p<0.01和**** p <0.0001。
3.4. TG甲型流感病毒的翻译后阻断可在致命病毒攻击中保护小鼠
我们先前发现TG抑制NHBE细胞和NPTr细胞中流感病毒复制的过程中,病毒NP和M1蛋白的病毒转录或产量几乎没有或没有变化,表明该病毒在翻译后被阻断了[14]。由于病毒NP和M1蛋白在胞质核糖体上被翻译成核输入,因此我们在这里检查了通过ER-高尔基体运往宿主细胞膜的病毒蛋白(HA,NA和M2)[25]。来自TG引发的NPTr细胞的病毒蛋白分析(图7A,B)显示,所有其他病毒蛋白的表达似乎也未受影响,这表明TG差异性靶向了流感病毒,RSV和冠状病毒的复制周期。由于TG的抗病毒作用在酸性pH值而不是在碱性pH值下稳定(图7C,D),因此在小鼠致命流感病毒攻击中评估了TG的口服治疗(感染后)功效。组中的每只BALB / c小鼠(每组n = 8)首先经鼻内感染3种MLD50的PR8 / H1N1病毒,第二天每天一次通过管饲法给予TG或oseltamivir,持续5天。低口服剂量TG(根据经验选择1.5 µg / kg /天)所赋予的保护作用与高剂量奥司他韦(45 mg / kg /天)对小鼠的保护作用相似。在治疗上,与PBS-PBS相比,TG治疗组的存活率显着提高(图7E),感染后3天和5天(dpi)的病毒脱落减少(图7F),体重减轻程度较轻(图7G,H)。 DMSO控制。七只,八只和两只小鼠全部在TG,奥司他韦和PBS +DMSO组中存活。在所有三个参数中,用TG和奥司他韦治疗的小鼠之间没有显着差异。因此,口服TG在遭受致死性流感病毒攻击的小鼠中赋予治疗保护。
图7. TG在翻译后抑制甲型流感病毒,对酸稳定,在致命病毒攻击中具有治疗性保护小鼠的作用。 (A)由TG引发的NPTr细胞引起的流感后代产量急剧下降是(B),同时病毒蛋白没有下降。将细胞用TG灌注30分钟,用PBS洗涤两次,并以0.5 MOI的浓度感染USSR病毒2小时。之后,将细胞再次用PBS洗涤3次,并在无血清培养基中温育24小时,该培养基中添加了0.2 µgl /mL的TPCK胰蛋白酶。(A)用相应的培养基样品进行聚焦形成测定,以确定活病毒的产量(ffu / µL)。所示的显着性基于单向ANOVA(Dunnett的多重比较)和相对于相应DMSO对照的存活子代百分比。 (B)病毒蛋白,包括通过ER-高尔基体处理的蛋白(HA,NA和M2),未显示TG引发的细胞减少。(C,D)TG的抗病毒活性在酸性但不是碱性条件下是稳定的。(C)首先,将所用的TG首先按照指示在pH 1.5(在30mM盐酸中)孵育不同的时间,并用氢氧化钠中和,然后以0.5 µM的终浓度应用于细胞中30分钟。 (D)首先将所用的TG在pH12.0(10 mM氢氧化钠)中孵育2小时,并用盐酸中和,然后以0.5 µM的最终浓度将其应用于细胞30分钟。在以0.5 MOI的感染率感染苏联H1N1病毒后二十四小时,将感染的培养基用于6小时聚焦形成试验,以免疫检测病毒NP以确定子代病毒的产量(ffu/ µL)。除非另有说明,否则重要性基于单向ANOVA Tukey的多重比较,并且病毒减少的百分比与相应的DMSO控件有关。 (E–H)TG在小鼠致命流感病毒攻击中的治疗保护。组中的每只BALB / c小鼠(每组n= 8)都经鼻内感染了3种MLD50的PR8 / H1N1病毒。然后每天给每只小鼠口服一次TG(1.5 µg / kg /天),奥司他韦(45mg /kg /天)或PBS + DMSO,持续5天;第一次剂量为12 hpi。在14d内记录生存率(E),通过TCID50分析的肺病毒滴度(F)和体重变化(G,H)。每个时间点代表平均值±SEM。 Kaplan–Meier方法用于生存分析。相对于相应的TG组显示了显着性。 * p <0.05,** p <0.01,*** p <0.001和****p<0.0001。
点击:查看文章结论
查看更多冠状病毒文章
查看更多医学文章
试用免费文档翻译功能
免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。
来源于:mdpi
2021-02-24 16:25:25

Thapsigargin是主要人类呼吸道病毒的广谱抑制剂:冠状病毒,呼吸道合胞病毒和A型流感病毒查看前部分内容查看文献图集4. 讨论 我们最近报道,TG在非细胞毒性水平下诱导了有效的宿主抗病毒反应,从而阻断了甲型流感病毒的复制[14]。在这里,我们进一步证明了TG是一种酸稳定的广谱抑制剂,对RSV,冠状病毒(OC43和SARS-CoV-2)和A型流感病毒非常有效。三种主要的RNA病毒类型代表不同的病毒基因组组成和生命周期。在此,TG的抗病毒效力是对可逆的ER应激激活的生理反应,与早期实验使用TG(长时间以高µM浓度)作为凋亡诱导剂的实验形成鲜明对比,后者是最终阶段ER应激的结果。迄今所有积累的证据表明,TG的抗病毒方案无细胞毒性,对RSV,OC43病毒和流感病毒显示出高选择性,并且在小鼠中没有任何明显的不良影响。先前的工作没有发现在连续的次优TG存在下,没有由甲型流感病毒的细胞传代引起的病毒抗性[14]。短TG暴露后至少48小时的持久抗病毒状态,以及其在主动感染之前或期间抑制不同病毒的能力,使得TG(或其衍生物)作为预防性和治疗性抗病毒候选物具有吸引力。此外,TG的抗病毒性能在分别抑制冠状病毒OC43和RSV方面明显优于RDV和利巴韦林。 TG作为抗病毒药的另一个优势是,它在单独的感染以及共同感染中对冠状病毒(OC43和SARS-CoV-2)和流感病毒(苏联H1N1和pdm H1N1)的抑制作用相同。流行性感冒管理的主要问题之一是需要在鼓动治疗之前通过实验室或快速诊断测试确认感染。或根据经验进行治疗,并可能使用针对非流感病原体的流感特异性疗法。诸如TG之类的广谱抗病毒药可能会及时改变这种范例。为了控制高传染性疾病,必须使用有效的疫苗和抗病毒药。在当前的COVID-19大流行中,疫苗已开始可用于大规模疫苗接种。但是,在一般人群中,尚无针对COVID-19的批准用于预防或早期感染社区的抗病毒药。 WHO团结试验[26]没有发现所检查的抗病毒药物(RDV,HC和IFNB1a)对死亡率或住院时间有任何重大影响。当前用于治疗COVID-19的其他药物,例如地塞米松和氢化可的松,是有症状的免疫调节剂,可调节病毒诱导的“细胞因子风暴”的破坏作用,而不是抗病毒药。因此,即使在有疫苗的情况下,也非常需要可以在活动性感染期间,尤其是在疾病的初始阶段进行干预的有效抗病毒药物。鉴于冠状病毒突变体的出现可能会破坏目前正在使用和开发中的疫苗,以及在人群中共同流行的其他主要呼吸道病毒,因此,对于区分当前和未来的大流行,更需要有效的抗病毒药物,不能区分病毒类型。如果可以将这种口服的广谱抗病毒药分配用于早期干预,而无需建立具体的呼吸道病毒诊断方法,那么在一般实践中,可以预料到TG比目前的抗病毒药具有巨大的潜在实用价值。这种抗病毒药将在减少SARS-CoV-2和其他呼吸道病毒的公共卫生负担方面与疫苗起不可或缺的作用。我们先前已经确定,TG诱导的ER应激导致UPR是关键的固有免疫驱动程序,其介导一系列以宿主为中心的抗病毒过程,以阻断A型流感病毒复制[14]。在最近的预印本中,据报道,TG在低于其细胞毒性范围的浓度下可有效抑制冠状病毒(hCoV-229E,MERS-CoV,SARS-CoV-2)复制[27]。然而,由原发性内质网应激反应引起的机制性抗病毒基础仍有待充分阐明。值得注意的是,TG处理触发了通用宿主抗病毒防御的下游频谱,这些防御在阻止不同RNA病毒的复制方面非常有效。 TG引发始终增加ER应激基因(DDIT3,HSPA5和HSP90B1)呈剂量依赖性。感染了RSV和OC43病毒TG引发的细胞进一步升高了ER应激基因的表达,但减弱了ER应激基因的诱导。RIG-I相关基因。对于两种病毒类型,TG对子代病毒产生的抑制作用都伴随着病毒转录和病毒蛋白表达的降低。相反,如所检查的所有病毒蛋白检测均无变化所证明,TG靶向翻译后的流感病毒复制,可能涉及翻译后病毒蛋白的修饰和转运。TG引发的细胞感染流感病毒也减弱了ER应激基因反应,但增强了RIG-I相关基因的表达[14]。总之,TG在病毒复制的不同阶段针对呼吸道病毒。 RSV和冠状病毒OC43的转录水平受到抑制,而甲型流感病毒则被翻译后靶向。 宿主ER应激和RIG-I相关反应的差异,以及RSV和OC43病毒感染以及TG引发细胞中的流感病毒在病毒阶段抑制方面的差异,表明在激活的多个抗病毒过程中存在功能冗余通过TG在生命周期的不同阶段控制不同的病毒。因此,在TG启动的细胞中,对感染的转录ER应激反应是病毒依赖性的。 RSV和OC43病毒感染在TG引发的细胞中引起强烈的ER应激基因表达,这又与RIG-I相关的转录激活减弱有关。相反,流感病毒感染减弱了TG引发细胞中ER应激基因的转录激活,这与RIG-I相关的转录激活升高有关。值得注意的是,功能性I型干扰素系统似乎在TG-ER应激介导的SARS-CoV-2复制和甲型流感病毒复制的抑制中至关重要[14]。鉴于在TG引发的细胞中,RIG-I相关的应答在RNA病毒类型之间有所不同,因此这种抗病毒信号通路的定性应答(如时间调节)可能比仅仅抑制特定病毒的应答强度更重要。复制。无论ER应激对感染的反应方式如何,TG显然对所有三种主要病毒类型均具有高度保护作用。总之,我们认为TG(或其衍生物)是一种有前途的口服活性广谱抗SARS-CoV-2,RSV和流感病毒的抗病毒药,有望成为下一轮X病大流行的防御者。 2. 专利权PCT / GB2019 / 050977和PCT / GB2020 / 052479涵盖了TG和其他与结构相关的化合物在抗病毒治疗中的用途。 作者贡献:S.A.-B.,C.A.P。 L.V.G .:概念化,调查,形式分析,撰写-原始草案的准备,审查和编辑; J.J.,J.P.,Z.J.,P.S.,B.L.W。 J.Y .:调查,形式分析,审查和编辑; A.C.B.,K.H.M.,P.G.,C.J.H.,J.N.-V.-T.,I.H.B。和J.L .:概念化,形式分析,审查和编辑; K.-C.C .:概念化,调查,形式分析,资金筹集,撰写-原始草案准备,审查和编辑。所有作者均已阅读并同意该手稿的发行版本。 资金来源:SAB在埃及高等教育和科学研究部博士奖学金上; CAP由生物技术和生物科学研究委员会(英国)博士培训计划2(BB /M008770/1)资助;这项工作部分由医学研究理事会概念信任计划(MC_PC_18058),诺丁汉大学,中国国家重点研究与发展计划(2016YFD0500204)和英国环境,食品与农村事务部(DEFRA)资助。 )以及下放的苏格兰和威尔士政府,授权号SV3700;资助者在本文的研究设计,分析,解释或撰写中不起作用。 SARs-CoV2病毒分离物是通过欧洲病毒库全球(EVA-GLOBAL)项目获得的,该项目已从欧盟的2020研究与创新计划中获得了资助,资助项目编号为871029。 利益冲突:J.N.-V.-T。目前已借调到英格兰卫生与社会保健部(DHSC)。本手稿表达的观点是作者的观点,不一定是DHSC的观点。其余作者声明没有利益冲突。 参考1. 中心。;郭浩;周鹏;施志霖SARS-CoV-2和COVID-19的特征。纳特微生物牧师。2020年。[CrossRef] [PubMed]2. I.D.A. Santos; Grosche,V.R .; F.R.G.的Bergamini; Sabino-Silva,R .;雅迪姆(A.C.G.)抗冠状病毒的抗病毒药:用于SARS-CoV-2治疗的候选药物?正面。微生物。 2020,11,1818。[CrossRef] [PubMed]3. 对手。;袁S.尹X.马丁·桑乔N.松永;帕西湖;Burgstaller-Muehlbacher,S。 De Jesus,P.D .; Teriete,P .;赫尔,M.V .;等。通过大规模的复合用途发现SARS-CoV-2抗病毒药物。自然2020,586,113–119。 [CrossRef] [PubMed]4. 丽娜(B.) C. Boucher;奥斯特豪斯A.S. Monto; M. R.J.惠特利; Nguyen-Van-Tam,J.S.甲型流感感染信息研究中的五年监测奥司他韦耐药性在甲型流感感染患者中的出现。流感其他呼吸道。病毒2018,12,267–278。 [CrossRef] [PubMed]5. 侯赛因,M。加尔文(H.D.); aw,T.Y .;纳特福德(Nutsford) Husain,M.甲型流感病毒的耐药性:流行病学和管理。感染。抗药性。 2017,10,121–134。 [CrossRef]6. 今井M.山下; Y.酒井田川Iwatsuki-Horimoto,K .; M.木曾;村上,J. A. Yasuhara;高田伊藤市中岛(Nakajima);等。从日本患者身上分离出来的对baloxavir敏感度降低的A型流感病毒变异体很适合并通过呼吸道飞沫传播。纳特微生物。 2020,5,27-33。 [CrossRef] [PubMed]7. 戈德希尔(D.H.); te Velthuis,A.J.W .; R.A.Fletcher; Langat,P .; M.Zambon;拉肯比(A. W.S.巴克莱流感病毒对favipiravir的耐药机制。进程Natl。学院科学美国,2018,115,11613–11618。 [CrossRef]8. 坦佩雷佩特克(A.萨拉塔角沃纳(Wallner) T.库尔迈斯特; Cazares-Körner,A .; T.Visnes;马里兰州赫塞尔曼; E. Kunold; E. Wiita;等。针对病原性RNA病毒复制所必需的宿主因子的新型广谱抗病毒抑制剂。病毒2020,12,1423。[CrossRef] [PubMed]9. 克拉默,F .;史密斯(G.J.D.); R.A.M. Fouchier; Peiris,M .; Kedzierska,K .;多尔蒂(P.C);Palese,P.;肖,M.L .; Treanor,J .; Webster,R.G .;等。流感。纳特版本号Dis。底漆2018,4,3. [CrossRef]10. 战斗,工商管理硕士;麦克莱伦(J.S.呼吸道合胞病毒进入以及如何阻止它。纳特微生物牧师。 2019,17,233–245。 [CrossRef]11. Collins,P.L .;费恩斯河;格雷厄姆(BS)呼吸道合胞病毒:病毒学,反向遗传学和疾病的发病机理。 Curr。最佳。微生物。免疫2013,372,3–38。 [考研]12. 特纳(T.L.); B.T. Kopp;保罗(Paul)兰德格雷夫小Hayes,D .;Thompson,R.呼吸道合胞病毒:当前和新兴的治疗选择。临床结果结果。 2014,6,217–225。 [CrossRef] [PubMed]13. 利顿(J.韦斯特林,M。Hanley,M.R.Thapsigargin抑制钙泵的肌浆网或内质网Ca-ATPase家族。 J.Biol。化学1991,266,17067-17071。 [CrossRef]14. 古尔德(L.V.);杨建江Z.张华带领。;埃默斯(R.D.); Dottorini,T .;蒲建刘建张凯毒胡萝卜素在无细胞毒性水平下会诱导有效的宿主抗病毒反应,从而阻断甲型流感病毒的复制。病毒2020,12,1093。[CrossRef]15. 法拉利,M。 A.Scalvini;明尼苏达州洛西奥; A. Corradi;桑奇尼,M。 E.Bignotti; E. Milanesi;阿杰蒙·马桑(Ajmone-Marsan); Barlati,S。贝洛蒂(D.等。建立和表征用于病毒学诊断实验室的两种新的猪细胞系。J.维罗尔方法2003,107,205–212。[CrossRef]16. P.A. Jorquera;特里普(RA)通过噬菌斑测定和免疫染色测定定量RSV感染颗粒。在人类呼吸道合胞病毒中:方法和协议;拉皮(Ripp),拉科(Jorquera),拉美(P.A.),编辑;施普林格:2016年,美国纽约,纽约;第33–40页。17. Spearman,C。没有高斯公式的对与错情况(恒定刺激)的方法。 Br。 J. Psychol。 1908,2,227。[CrossRef]18. Kärber,G.对药理学系列实验的集体治疗做出了贡献。 Naunyn-Schmiedeberg的Arch。Exp。Pathol。 Pharmacol。 1931,162,480-483。19. 里德,L.J .;Muench,H。一种估算50%终点的简单方法。是。 J.流行病。1938,27,493–497。 [CrossRef]20. Mirabelli,C .;贾斯珀斯(M. Boon,M .; Jorissen,M .; M.库克尼;巴迪奥特(D.) Chaltin,P .; A. Marchand;Neyts,J .; Jochmans,D.人呼吸道上皮细胞中呼吸道合胞病毒(RSV)抑制剂的差异抗病毒活性。 J.抗微生物剂。化学药剂师。 2018,73,1823–1829。 [CrossRef]21. A.R.Fehr;Perlman,S.冠状病毒:其复制和发病机理的概述。方法生物学2015,1282,1–23。22. E. Keyaerts;李珊;维杰根湖; Rysman,E .; Verbeeck,J .;范·兰斯特(M. Maes,P.氯喹对新生小鼠中人类冠状病毒OC43感染的抗病毒活性。抗微生物剂。代理商Chemother。 2009,53,3416。[CrossRef]23. 刘建曹河;徐敏王X.张华胡华;李Y胡中;钟,W。 Wang,M.羟氯喹,一种毒性较小的氯喹衍生物,在体外可有效抑制SARS-CoV-2感染。 Cell Discov。 2020,6,16,[CrossRef] [PubMed]24. 布朗(A.J.);元俊杰;格雷厄姆(R.L.);丁农(K.H.) A.C. Sims;冯建元; Cihlar,T .;马萨诸塞州丹尼森; R.S. Baric; T.P. Sheahan广谱抗病毒瑞姆斯韦以高度依赖于RNA的RNA聚合酶抑制人类地方性和人畜共患的三角冠状病毒。抗病毒Res。 2019,169,104541.[CrossRef] [PubMed]25. 窦D.革命,河。 Östbye,H。王华; Daniels,R.甲型流感病毒细胞的进入,复制,病毒体组装和移动。正面。免疫2018,9,1581。[CrossRef] [PubMed]26. 潘华;佩托河;卡里姆(Q.A.)亚历杭德里亚,M。 Henao-Restrepo,A.M .;加西亚(C.H.) Kieny,M.-P .; R.Malekzadeh;Murthy,S .; Preziosi,M.-P .;等。针对COVID-19的重新定向的抗病毒药物– WHO WHO SOOLIDARITY中期试验结果。 medRxiv2020。[CrossRef]27. 麻省沙班; C.Müller; Mayr-Buro,C .;韦瑟,H。阿尔伯特(B.V.); Weber,答:美国Linne;海恩(Hain)巴巴耶夫北卡罗来纳州;等。通过化学内质网应激抑制培养细胞中的冠状病毒复制。bioRxiv2020。[CrossRef]点击:查看相关冠状病毒文章 查看相关医学文章 使用福昕翻译功能免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:mdpi
2021-02-24 16:25:07

控制大流行病需要采取预防病毒传播的措施,但这一功能很难衡量。
可以阻止病毒传播的疫苗将有助于控制大流行。图片来源:Andrea Fasani / EPA-EFE / Shutterstock
随着各国推出预防COVID-19的疫苗,正在进行研究以确定注射疫苗是否还可以阻止人们感染和传播SARS-CoV-2病毒。如果将疫苗传播给足够多的人,则可以帮助控制该流行病。
初步分析表明,至少某些疫苗可能具有传播阻断作用。但是,要确认这种影响以及这种影响的强度是很棘手的,因为给定区域感染的下降可能是由其他因素(例如封锁和行为改变)解释的。不仅如此,该病毒还可以从无症状携带者传播,这使得很难检测到这些感染。
“这些是最难的研究之一,”马萨诸塞州波士顿的哈佛大学陈陈公共卫生学院传染病流行病学专家马克·利普西奇说。他说:“我们所有人都在那里,不停地尝试查看从确实出现的少量数据中可以得到什么。” 预计未来几周将获得一些研究的结果。
停止感染?
尽管大多数COVID-19疫苗的临床试验都表明疫苗可以预防这种疾病,但一些试验结果还提供了一些线索,表明注射
2021-02-22 19:36:27

由 纽约大学
PNG / CC0公共领域
SARS-CoV-2突触蛋白的突变是英国,南非和巴西出现的相关变体中的几种遗传突变之一,使该病毒在人细胞中的传染性比普通人高出八倍。根据发表在《eLife》杂志上的研究,该病毒起源于中国。
由纽约大学,纽约基因组中心和西奈山的研究人员领导的这项研究证实了D614G突变使SARS-CoV-2更易于传播的发现。
纽约大学生物学助理教授内维尔·桑贾纳(Neville Sanjana)表示:“自我们最初进行这项研究以来的几个月中,D614G突变的重要性不断提高:该突变已接近普遍流行,并已包括在所有当前关注的变异中。纽约大学格罗斯曼医学院神经科学与生理学教授,纽约基因组中心核心教员。“确认这种突变导致更多的可传播性,可能在一定程度上解释了为什么病毒在过去的一年中如此迅速地传播。”
SARS-CoV-2穗状蛋白中的D614G突变(通常称为“ G变体”)可能在2020年初出现,现在是全美国SARS-CoV-2病毒中最流行和最主要的形式在全球许多国家 随着多个突变的传播,研究人员一直在努力了解这些突变的功能意义,以及它们是否有意义地改变了病毒的传
2021-02-18 19:39:28

剑桥大学 自1901年以来,由于气候变化导致地理范围的变化,蝙蝠的本地数量有所增加。放大的区域代表了SARS-CoV-2蝙蝠起源祖先的可能空间起源。图片来源:Robert Beyer博士 今天发表在《科学总数》杂志上的一项新研究,环境为气候变化可能在SARS-CoV-2(导致COVID-19大流行的病毒)的出现中发挥了直接作用的机制提供了第一个证据。 这项研究表明,在过去的一个世纪中,云南南部以及缅甸和老挝的邻近地区的植被类型发生了大规模变化。气候变化,包括温度升高,日照增加和大气中的二氧化碳(影响植物和树木的生长),已将自然栖息地从热带灌木地变为热带稀树草原和落叶林。这为主要生活在森林中的许多蝙蝠物种创造了合适的环境。 一个地区中冠状病毒的数量与存在的不同蝙蝠物种的数量紧密相关。该研究发现,在过去的一个世纪中,另外40种蝙蝠物种已迁入中国云南南部,其中藏有大约100多种蝙蝠传播的冠状病毒。这个“全球热点”是遗传数据表明可能出现SARS-CoV-2的区域。 研究人员罗伯特·拜尔(Robert Beyer)博士说:“上个世纪的气候变化使云南南部的栖息地更适合
2021-02-07 16:35:10

SARS-CoV-2穗蛋白在人类宿主细胞中激发细胞信号转导:对COVID-19疫苗可能后果的暗示通过铃木雄一郎和塞尔吉·吉奇卡(Sergiy G.Gychka) 1 美国华盛顿哥伦比亚特区乔治敦大学医学中心药理生理学系,美国200072 Bogomolets国立医科大学病理解剖学N2系,01601基辅,乌克兰;* 应与之联系的作者。收到:2020年12月15日接受:2021年1月8日发行时间:2021年1月11日 摘要:世界正遭受由严重急性呼吸系统综合症冠状病毒2(SARS-CoV-2)引起的2019年冠状病毒疾病(COVID-19)大流行。 SARS-CoV-2使用其刺突蛋白进入宿主细胞。目前正在开发将刺突蛋白引入人体内以诱发病毒中和抗体的疫苗。在本文中,我们注意到人类宿主细胞对刺突蛋白敏感地应答以引发细胞信号传导。因此,重要的是要意识到新的COVID-19疫苗产生的刺突蛋白也可能影响宿主细胞。我们应该仔细监测这些疫苗的长期后果,尤其是当将它们接种给其他健康个体时。有必要进一步研究SARS-CoV-2刺突蛋白对人细胞的影响以及适当的实验动物模型。关键词:细胞信号转导;病毒;肺炎; SARS-CoV-2;刺突蛋白疫苗 1. 介绍世界正遭受由严重的急性呼吸综合症冠状病毒2(SARS-CoV-2)引起的2019年冠状病毒大流行(SARS-CoV-2),这是一种正向单链RNA病毒[1,2]。截至2020年12月,全球有8000万人感染了SARS-CoV-2,造成180万人死亡。 SARS-CoV-2使用其病毒膜融合蛋白(称为刺突蛋白)结合血管紧张素转化酶2(ACE2)作为“受体”,以进入人类宿主细胞[3,4],引起严重的肺炎和急性呼吸窘迫综合征(ARDS)[5]。患有心血管疾病的老年患者特别容易出现严重的COVID-19病状,在某些情况下甚至会导致死亡,而年轻健康的人对产生严重症状的抵抗力很大[1,6,7]。随着COVID-19继续造成严重的健康,经济和社会问题,全世界都在等待有效疫苗的广泛推广,以结束这种流行病。I类病毒融合蛋白SARS-CoV-2峰值蛋白对于启动病毒与宿主细胞表面受体之间的相互作用,通过协助病毒与宿主细胞融合促进病毒进入宿主细胞至关重要膜。该蛋白由两个亚基组成:包含ACE2受体结合域(RBD)的亚基1(S1)和在融合过程中起作用的亚基2(S2)[3,4](图1)。 SARS-CoV-2刺突蛋白是开发COVID-19疫苗的主要目标。图1. SARS-CoV-2刺突蛋白的结构。刺突蛋白由亚基1(S1)和亚基2(S2)组成。 S1亚基包含与宿主细胞膜ACE2结合的受体结合域(RBD)。 S2亚基负责融合。在我们先前在第3节和第5节中描述的研究中,我们使用了全长S1(Val16-Gln690),该区域描绘了蓝色和红色区域,SARS-CoV-2的红色显示了仅含RBD的蛋白质(Arg319-Phe541)。刺突蛋白(GenBank登录号:QHD43416.1)。2. 基于穗蛋白的COVID-19疫苗的开发2020年COVID-19疫苗和治疗剂的快速发展归功于政府与私营部门之间的有效合作。2020年11月9日,辉瑞和BioNTech宣布他们的基于mRNA的候选疫苗BNT162b2对COVID-19的有效性超过90%[8]。这是令人欢迎的消息,因为它表明有效的疫苗可能很快就会出现。BNT162b2编码SARS-CoV-2突突蛋白以诱导病毒中和抗体[9,10]。更具体地说,它编码SARS-CoV-2的全长刺突蛋白,其中两个氨基酸在S2亚基中突变为脯氨酸以维持预融合构象,而其姊妹疫苗BNT162b1(同样来自辉瑞公司/ BioNTech)仅编码RBD SARS-CoV-2穗蛋白的结构,通过添加T4纤维蛋白折叠域来三聚化[9-11]。临床试验表明,BNT162b1 [11]和BNT162b12[9,10]均未显示严重的短期不良反应。2020年12月10日,发表了一项BNT162b大型临床试验的结果,表明该疫苗对16岁或16岁以上的人群提供了95%的保护[12]。但是,这些疫苗的长期后果尚不清楚。另一种有前途的疫苗,Moderna的mRNA-1273也是一种RNA疫苗,可编码全长SARS-CoV-2穗蛋白[13]。基于病毒载体的疫苗,例如阿斯利康(AstraZeneca)的AZD1222,它使用非复制性黑猩猩腺病毒载体[14],强生公司(Johnson&Johnson)的Ad26.COV2.S,非复制性腺病毒26系统[15]和Gam- Gamaleya流行病学和微生物研究所的COVID-Vac(Sputnik V)[16]均表达SARS-CoV-2刺突蛋白。 NVX-CoV2373(Novavax),一种基于蛋白质的重组疫苗[17],也是全长SARS-CoV-2穗蛋白。这些疫苗以及许多其他正在开发中的疫苗[18-20]将SARS-CoV-2刺突蛋白引入了我们的体内,从而刺激了抗体的产生和针对SARS-CoV-2的免疫力。3. SARS-CoV-2 Spike蛋白促进人类细胞中的细胞信号传导发现用重组SARS-CoV-2穗突蛋白S1亚基处理培养的原代人肺动脉平滑肌细胞(SMCs)或人肺动脉内皮细胞足以促进细胞信号转导,而无需其余病毒成分[ 21]。此外,我们对死于COVID-19的患者的死后肺组织的分析已确定这些患者表现出肺血管壁增厚,这是肺动脉高压(PAH)的标志[21]。基于这些结果,我们提出SARS-CoV-2突触蛋白(无其余病毒成分)触发细胞信号转导事件,可能促进肺血管重构和PAH以及其他心血管并发症[21,22]。在我们的细胞培养实验中,研究了两个都含有RBD的重组SARS-CoV-2刺突蛋白[21]。全长S1亚基蛋白包含大部分S1亚基(Val16–Gln690),而RBD S1亚基蛋白仅包含RBD区(Arg319–Phe541),如图1所示。用这些蛋白质处理肺动脉内皮细胞10分钟。我们发现,使用磷酸化特异性MEK抗体,单独的SARS-CoV-2全长S1亚基(浓度低至130 pM)激活了MEK,细胞外信号调节激酶(ERK)的激活剂和众所周知的细胞生长信号转导机制[23]。相比之下,在大鼠肺动脉SMC中,这种由刺突蛋白引起的细胞信号激活并未发生[21]。尽管现在众所周知ACE2是SARS-CoV-2突突蛋白与人宿主细胞结合的“受体”,以促进膜融合和获得病毒进入,但是ACE2的通常生理功能并不充当膜受体转导细胞内信号。 ACE2是一种I型整合膜蛋白,起羧肽酶的作用,将血管紧张素II裂解为血管紧张素(1-7)并调节血压[24,25](图2)。然而,十年前,Chen等。[26]报告了有趣的发现,表明ACE2充当细胞信号转导的膜受体,以响应SARS-CoV的突增蛋白(现在也称为SARS-CoV-1,该病毒在2002年引起SARS爆发) 2004)在人肺泡上皮细胞系A549中。SARS-CoV-1的刺突蛋白与SARS-CoV-2的刺突蛋白具有76–78%的同一性[27]。在他们的研究中,表明全长刺突蛋白与ACE2的结合触发了酪蛋白激酶II依赖性激活蛋白1(AP-1)转录因子的激活以及随后的基因转录事件[26]。他们在SARS-CoV-1 [26]和我们在SARS-CoV-2 [21]上的发现表明,刺突蛋白将ACE2(通常是肽酶)功能性地转化为膜受体,从而利用刺突将细胞信号转导。蛋白质作为其激活的配体(图2)。图2. ACE2的生物学功能。在生理情况下,ACE2充当羧肽酶,通过裂解苯丙氨酸(Phe)催化血管紧张素II(Ang II)水解为Ang(1-7)。在刺突蛋白的存在下,该酶成为细胞信号转导的膜受体,该信号使用刺突蛋白作为其激活的配体。库巴等。[28]表明,给小鼠注射重组SARS-CoV-1突突蛋白会降低ACE2的表达,并加剧酸诱导的肺损伤。在患有酸诱导的肺损伤的小鼠中,重组SARS-CoV-1刺突蛋白显着增加了血管紧张素II,而血管紧张素受体抑制剂氯沙坦减弱了刺突蛋白诱导的肺损伤的增强[28]。因此,这些体内研究表明,SARS-CoV-1的突触蛋白(无其余病毒)会降低ACE2的表达,增加血管紧张素II的水平,并加剧肺损伤。Patra等人还显示,不含其余病毒成分的SARS-CoV-2突突蛋白可激活细胞信号传导。 [29]。作者报告说,在人肺泡上皮细胞系A549或人肝上皮细胞系Huh7.5中通过瞬时转染的方式表达了全长SARS-CoV-2突突蛋白,并激活了NF-κB和AP-1转录因子以及p38和ERK丝裂原激活的蛋白激酶,释放白介素6。发现该细胞信号转导级联由下调ACE2蛋白表达的SARS-CoV-2突突蛋白触发,随后激活了1型血管紧张素II受体[29]。这些使用瞬时转染的实验可能反映了可能由基于RNA和病毒载体的疫苗触发的刺突蛋白的细胞内效应。这些结果共同强化了这样一种观念,即通过细胞信号转导的激活,人类细胞受到细胞外和/或细胞内刺突蛋白的敏感影响。4. 肺动脉高压PAH是一种无法治愈的严重疾病,可能会影响任何年龄段的男性和女性,包括儿童。 PAH中增加的肺血管阻力会导致右心衰竭并随后死亡。如果不进行治疗,被诊断为PAH的患者从诊断之时起平均只能存活2-3年[30,31]。即使采用目前可用的疗法,PAH患者中也只有60-70%可以存活三年[32-35]。 PAH难以检测,因为其症状(例如呼吸急促,疲劳和头晕)与其他常见的无生命危险的症状相似,并且必须通过有创右心导管检查来对PAH进行官方诊断[36] 。内皮功能障碍是PAH和COVID-19患者的共同特征[37,38]。多环芳烃的“爆发”与某些药物或毒素的暴露有关[39]。 PAH的主要爆发发生在1965年,并与一种减肥瘦身药aminorex有关[39,40]。服用这种药物的人中约有0.2%会发展为PAH [40]。引入氨甲x呤两年后就发现了一种流行病,这种流行病发生十年后,有一半的患者死亡[39]。我们研究了死于ARDS的COVID-19患者和H1N1流感感染患者的肺血管[21]。死后COVID-19患者肺部的肺动脉始终表现出血管壁增厚的组织学特征,这主要是由于中膜肥大所致。详细的病理学分析显示,血管与周围肺实质之间的边界变得不清晰,动脉中层内膜的SMC增大,SMC的细胞核肿胀,并且在SMC的细胞质中产生了液泡[21]。 ]。形态计量学分析确定中位肺血管壁厚度对于COVID-19患者,该值是15.4 µm,对于流感患者,该值是6.7µm,这些值彼此之间存在显着差异[21]。肺血管壁在胸部计算机X线断层扫描中也观察到了COVID-19患者的乳腺增厚[41,42]。因此,这些结果共同表明COVID-19与肺血管壁增厚有关。有必要对这种肺血管壁增厚是否与临床上显着的PAH相关以及是否存在突波蛋白在PAH发病机理中的作用进行研究。 5. 仅包含RBD的SARS-CoV-2突突蛋白不能诱导人类细胞中的细胞信号传导与全长刺突蛋白[26,29]或全长SARS-CoV-2刺突蛋白S1亚基[21]相反,我们发现仅含RBD的蛋白(图1)不会促进细胞信号传导。我们监测MEK活化的Western印迹结果表明,SEM磷酸化的MEK与MEK蛋白的平均比率值为0.05 0.003(未处理),1.90.07(使用全长S1蛋白处理)和0.05 0.003(仅使用RBD处理-含蛋白质)用于人肺动脉SMC;对于人肺动脉内皮细胞而言,分别为0.09 0.006(未处理),0.90 0.06(使用全长S1蛋白处理)和0.10 0.003(仅包含RBD的蛋白处理)[21]。 考虑到BNT162b2和许多其他COVID-19疫苗表达全长刺突蛋白,而仅包含S1和RBD的全长蛋白的不同作用可能很重要,而BNT162b1疫苗仅编码RBD区域[9-20] 。还有其他一些基于RBD的COVID-19疫苗也正在开发中[43]。基于RBD的疫苗可能免疫原性较低,但可能不会影响宿主细胞。因此,考虑到潜在的长期不利影响,他们的风险可能较小。但是,在上述SARS-CoV-1穗蛋白的体内研究中[28],仅包含RBD的缺失突变体也像全长穗蛋白一样使酸诱导的肺衰竭恶化。因此,需要进一步的工作来了解全长刺突蛋白和仅含RBD的蛋白在各种生物过程中的作用。6. 讨论区通常认为,病毒膜融合蛋白的唯一功能是允许病毒结合宿主细胞,以病毒进入细胞,从而释放遗传物质,并进行病毒复制和扩增。地点。但是,最近的观察表明,SARS-CoV-2突突蛋白本身可以触发细胞信号传导,从而导致各种生物学过程。可以合理地假设,在某些情况下,此类事件会导致某些疾病的发病机理。我们的实验室仅测试了SARS-CoV-2峰值蛋白在肺血管细胞以及与PAH的发生有关的蛋白中的作用。但是,这种蛋白质也可能影响全身和冠状血管的细胞,引发其他心血管疾病,例如冠状动脉疾病,全身性高血压和中风。除心血管细胞外,其他表达ACE2的细胞也可能受到SARS-CoV-2峰值蛋白的影响,这可能会导致不良的病理事件。因此,重要的是要考虑由新的COVID-19疫苗产生的SARS-CoV-2突突蛋白触发某些人中促进PAH,其他心血管并发症和/或其他组织/器官并发症的细胞信号事件的可能性。(图3)。我们将需要仔细监控将刺突蛋白引入人体的COVID-19疫苗的长期后果。此外,尽管不会很快获得有关基于刺突蛋白的COVID-19疫苗可能产生的长期后果的人类数据,但必须尽快采用适当的实验动物模型以确保SARS-CoV-2棘突蛋白不会引起任何PAH发病机理或任何其他慢性病理状况的迹象。 图3. SARS-CoV-2刺突蛋白的可能作用。完整病毒的SARS-CoV-2突突蛋白靶向宿主细胞的ACE2,以促进膜融合和病毒进入。 SARS-CoV-2突突蛋白还可以在人细胞中引发细胞信号转导[21,29]。 COVID-19疫苗将刺突蛋白引入人体。除了引起抑制病毒进入的免疫反应外,COVID-19疫苗产生的突触蛋白还可能影响宿主细胞,可能引发不良事件。有必要针对此可能性进行进一步调查。 7. 结论总之,基于SARS-CoV-2穗蛋白的COVID-19疫苗开发的最新进展令人激动,并阐明了如何结束当前的大流行。如果这些疫苗没有表现出任何急性不良反应,它们应该使患有基础疾病的老年人受益。但是,我们需要仔细考虑它们的长期后果,尤其是当将它们用于其他健康的个体以及年轻人和儿童时。除了评估可从感染SARS-CoV-2的个体以及已接受基于穗状蛋白的疫苗的人获得的数据之外,进一步研究SARS-CoV-2穗状蛋白在人细胞和适当动物中的作用型号是保证的。 作者贡献:概念化,Y.J.S .;验证,Y.J.S。和S.G.G .; Y.J.S.调查和S.G.G .;资源,Y.J.S。和S.G.G.;写作-原始草稿,Y.J.S .;写作-审核和编辑,Y.J.S.和S.G.G.;可视化,Y.J.S .;监督,Y.J.S.; Y.J.S.项目管理;资金获取,Y.J.S.两位作者均已阅读并同意该手稿的发行版本。资金:这项研究由美国国立卫生研究院(NIH)资助,授权号R21AI142649,R03AG059554和R03AA026516,资金由Y.J.S.内容仅是作者的责任,并不一定代表NIH的官方观点。利益冲突:作者声明没有利益冲突。资助者在研究的设计中没有作用。在数据的收集,分析或解释中;在手稿的写作中;或决定发布结果。参考文献(展示部分,可至原网站查看全部1~43)1. 黄昌王Y;李旭任丽;赵建胡Y;张丽;范G;徐建顾旭等。中国武汉市2019年新型冠状病毒感染患者的临床特征。柳叶刀2020,395,497–506。 [CrossRef]2. 吴昌;陈旭蔡Y;夏J.周X.徐珊;黄辉;张丽;周X.杜C.等。中国武汉冠状病毒病2019肺炎患者与急性呼吸窘迫综合征和死亡相关的危险因素。 JAMA实习生。中2020年,e200994。 [CrossRef]3. 严河;张Y李Y霞郭Y; Zhou,Q.全长人ACE识别SARS-CoV-2的结构基础。科学2020,367,1444–1448。 [CrossRef]4. 戴,W。他,L。张旭。蒲建沃罗宁(D.江南周Y; Du,L.2019年新型冠状病毒的受体结合结构域(RBD)的表征:对RBD蛋白作为病毒附着抑制剂和疫苗的开发意义。细胞。大声笑免疫2020,17,613–620。 [CrossRef]5. 徐中施力;王Y;张建。黄丽;张成;刘珊;赵鹏;刘华;朱力;等。与急性呼吸窘迫综合征相关的COVID-19的病理发现。柳叶刀呼吸。中2020,8,420–422。 [CrossRef]6. 李宝杨建赵峰;智林;王X.刘力; Bi,Z .;赵燕。中国心血管代谢疾病的流行及其对COVID-19的影响。临床Res。乙二醇。 2020,109,531–538。 [CrossRef]7. 杨建郑Y苟X. Pu,K .;陈正郭庆;Ji,R .;王华;王Y; Zhou,Y .. SARS-CoV-2感染患者的合并症患病率及其影响:系统评价和荟萃分析。诠释J.感染。 Dis。2020,94,91–95。 [CrossRef]8. 辉瑞公司和BioNTech宣布针对COVID-19的疫苗候选者在第三阶段研究的首次中期分析中取得了成功。可在线获得:https://www.businesswire.com/news/home/20201109005539/en/(2020年11月9日访问)。9. 沃尔什(E.E.); Frenck,R .; Falsey,A.R.; N.基钦;阿布萨隆,J。 Gurtman,A .;洛克哈特。 Neuzil,K .; M.J. Mulligan;贝利河;等。选择基于RNA的COVID-19疫苗BNT162b2进行关键功效研究。medRxiv2020。[CrossRef]10. 沃尔什(E.E.);小弗伦克(R.W.)Falsey,A.R .; N.基钦;阿布萨隆,J。 Gurtman,A .;洛克哈特。 Neuzil,K .; M.J. Mulligan;贝利河;等。两种基于RNA的COVID-19候选疫苗的安全性和免疫原性。 N. Engl。 J. Med。2020,383,2439-2450。 [CrossRef]11. M.J. Mulligan;肯塔基州莱克; N.基钦;阿布萨隆,J。Gurtman,A .;洛克哈特。 Neuzil,K .; V. Raabe;贝利河;斯旺森(Kwan)等。成人COVID-19 RNA疫苗BNT162b1的I/ II期研究。自然2020,586,589–593。[CrossRef]12. F.P.波拉克;Thomas,S.J .; N.基钦;阿布萨隆,J。 Gurtman,A .;洛克哈特。佩雷斯J.L .;佩雷斯·马克(G. E.D. Moreira; Zerbini,C。等。 BNT162b2 mRNA COVID-19疫苗的安全性和有效性。 N. Engl。J.Med。 2020,383,2603–2615。(在新闻)。[CrossRef]13. 洛杉矶杰克逊; E.J.安德森; N.G.鲁法尔;罗伯茨(P.C.) Makhene,M。 R.N.麦卡洛(McCullough),硕士; Chappell,J.D.;马萨诸塞州丹尼森;史蒂文斯(L.J.);等。针对SARS-CoV-2的mRNA疫苗-初步报告。 N. Engl。 J. Med。 2020,383,1920–1931。 [CrossRef]14. 弗雷加蒂(P.M.);Ewer,K.J .;P.K. Aley; B.安格斯贝克尔,S。 Belij-Rammerstorfer,S .;贝拉米Bibi,S .; M.Bittaye; E.A. Clutterbuck;等。针对SARS-CoV-2的ChAdOx1 nCoV-19疫苗的安全性和免疫原性:一项1/2期,单盲,随机对照试验的初步报告。柳叶刀2020,396,467–478。 [CrossRef]15. N.B.梅尔卡多; Zahn,R .; Wegmann,F。 C.钱德拉舍卡(A.于俊刘建彼得·L;麦克马汉L.H. Tostanoski;等。单剂Ad26疫苗可预防猕猴中的SARS-CoV-2。自然2020,586,583–588。 [CrossRef]16. 罗格诺夫(D.Y.);伊利诺伊州多尔芝科娃; O.V.祖布科娃;杜克瓦图林(A.I.); Shcheblyakov,D.V .; Dzharullaeva,美国;格罗索娃(D.M.); A.S.埃罗霍娃; A.V. Kovyrshina; A.G. Botikov;等。基于rAd26和rAd5载体的异源初免加强型COVID-19疫苗的两种制剂的安全性和免疫原性:来自俄罗斯的两项开放式,非随机1/2期研究。柳叶刀2020,396,887–897。 [CrossRef]17. Guebre-Xabier,M .;北帕特尔;田建勋;周乙Maciejewski,S。 Lam,K .; AD Portnoff;麻萨诸塞州麻萨诸塞州;弗里曼(M.B.)彼德拉(Piedra)等。 NVX-CoV2373疫苗可保护猕猴的上呼吸道和下呼吸道免受SARS-CoV-2攻击。疫苗2020,38,7892–7896。 [CrossRef]18. Kaur,S.P .;Gupta,V. COVID-19疫苗:全面的状态报告。病毒库。 2020,288,198114。[CrossRef]19. 董瑜;戴T.魏Y;张丽;郑敏;Zhou,F。SARS-CoV-2候选疫苗的系统评价。信号传导。目标。那个2020,5,237。[CrossRef]20. Krammer,F. SARS-CoV-2疫苗正在开发中。自然2020,586,516–527。[CrossRef]21. 铃木(Y.J.);尼古拉连科(S.I.)弗吉尼亚州迪布罗娃; Y.V. Dibrova;瓦西里克(Vasyk)纽约州诺维科夫;N.V.舒尔特;Gychka,S.G。SARS-CoV-2在肺血管细胞中掺入蛋白介导的细胞信号传导。血管Pharmacol。 2020,106823,(在线印刷之前)。 [CrossRef]22. Suzuki,Y.J.COVID-19发病机理的病毒蛋白片段理论。中假设2020,144,110267。[CrossRef]23. 张伟刘浩MAPK信号通路在哺乳动物细胞中调节细胞增殖。细胞水库。 2002年12月9-18日。 [CrossRef]24. Gheblawi,M .;王K;维维罗斯(A.) Q.阮钟建特纳(A.J.); M.K. Raizada;格兰特(M.B.) Oudit,G.Y。血管紧张素转换酶2:SARS-CoV-2受体和肾素-血管紧张素系统的调节剂:庆祝ACE发现20周年。大约Res。 2020,126,1456-1474。 [CrossRef]25. 华纳(F.J.);史密斯(A.I.);胡珀(N.M.);特纳(A.J.血管紧张素转换酶2:分子和细胞的角度。细胞。大声笑生命科学2004,61,2704–2713。 [CrossRef]26. 陈P Chang,S.C.;吴慧妍;俞TC Wei,W.C .; Lin S.; Chien,C.L .;张M通过严重的急性呼吸综合症冠状病毒刺突-ACE2信号转导趋化因子(C-C主题)配体2。 J.维罗尔2010,84,7703–7712。 [CrossRef]点击:查看更多冠状病毒分类文章 查看更多医学分类文章 文档翻译功能免费试用免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:mdpi
2021-02-07 16:30:56
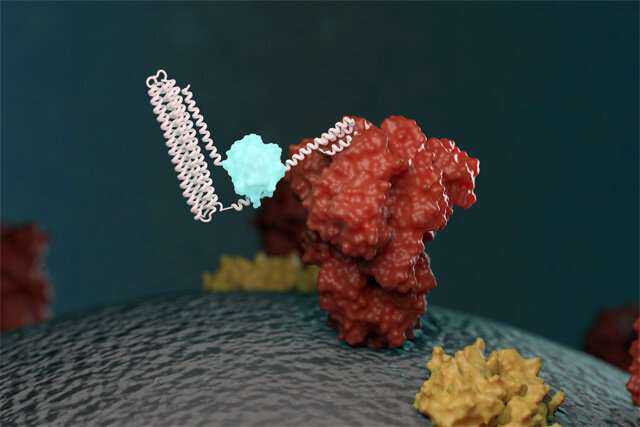
由 华盛顿大学新的生物传感器与目标分子结合并发光的插图。生物传感器的创建是由UW蛋白质设计医学研究所领导的。图片来源:Ian Haydon /西澳大学医学院的蛋白质设计科学家创造了一种新的方法来检测构成大流行冠状病毒的蛋白质以及针对大流行冠状病毒的抗体。他们设计了基于蛋白质的生物传感器,当与病毒成分或特定的COVID-19抗体混合时会发光。这一突破可以在不久的将来实现更快,更广泛的测试。该研究发表在《自然》上。今天,要诊断冠状病毒感染,大多数医学实验室都依赖一种称为RT-PCR的技术,该技术可放大病毒的遗传物质,以便可以看到它。此技术需要专门的人员和设备。它还消耗了全球范围内需求旺盛的实验室用品。供应链短缺使美国及其他地区的COVID-19测试结果放慢了速度。为了直接检测患者样本中的冠状病毒而不需要进行基因扩增,由生物化学教授,西澳大学医学院蛋白质设计研究所所长戴维·贝克(David Baker)带领的一组研究人员使用计算机来设计新的生物传感器。这些基于蛋白质的设备识别病毒表面上的特定分子,与它们结合,然后通过生化反应发光。抗体测试可以揭示一个人过去是否患有过COVID-19。它被用来追踪大流行的蔓延,但是它也需要复杂的实验室用品和设备。Alfredo Quijano-Rubio在西雅图的UW医学蛋白质设计研究所进行研究。他领导了设计和测试新型发光生物传感器以检测冠状病毒蛋白和抗体的工作。对该模型生物传感器的未来修改也可能对检测癌症和传染病中其他临床上重要的分子有用。图片来源:Ian Haydon / UW蛋白设计研究所威斯康星大学同一研究小组还创造了与COVID-19抗体混合后会发光的生物传感器。他们表明,这些传感器对可能也存在于血液中的其他抗体(包括针对其他病毒的抗体)不产生反应。这种敏感性对于避免假阳性测试结果很重要。贝克说:“我们已经在实验室中证明了这些新型传感器可以很容易地检测到模拟鼻液或捐赠的血清中的病毒蛋白或抗体。这项工作说明了从头进行蛋白质设计的能力,可以从头开始创建具有新的有用功能的分子装置。”除COVID-19以外,研究小组还表明,可以设计类似的生物传感器来检测医学上相关的人类蛋白质,例如Her2(某些形式的乳腺癌的生物标志物和治疗靶标)和Bcl-2(对淋巴瘤和某些癌症具有临床意义)。其他癌症),以及针对乙肝病毒的细菌毒素和抗体。 点击:查看更多化学文章 查看更多生物学文章 查看更多冠状病毒类文章 使用双语译文文档翻译功能 免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:phys
2021-01-31 17:15:05

人类冠状病毒:病毒与宿主相互作用的综述(上) 7.人冠状病毒与NF-κB途径NF-κB蛋白是转录因子家族,可调节基因表达以控制广泛的生物学过程,例如细胞死亡,炎症,先天性和适应性免疫反应。哺乳动物的NF-κB家族由五个成员组成,即RelA(也称为p65),RelB,c-Rel,NF-κB1p50和NF-κB2p52,它们在细胞质中形成二聚体。已经确定,病毒病原体经常靶向NF-κB途径以增强病毒复制,宿主细胞存活和宿主免疫逃避[199,200]。 NF-κB信号传导的主要途径有两种:经典途径和非经典途径。在经典途径中,潜在的NF-κB与抑制剂IκB蛋白形成复合物,并被隔离在细胞质中。如上所述,病毒病原体的存在会激活各种膜传感器,例如RIG-I,后者会诱导IκB激酶(IKK)复合物磷酸化IκB,并随后引起泛素化。 IKK复合物由三聚体亚基组成,包括两个催化亚基IKKα和IKKβ,以及调节亚基IKKγ(也称为NF-κB基本调节剂或NEMO)。因此,NF-κB从IκB的抑制作用中释放出来并转移到核中,在核中,它可以单独或与其他转录因子(包括AP-1,Ets和Stat)组合刺激靶基因的转录[201]。另一方面,非经典途径与IκB降解无关,而是依赖于可诱导的p100加工。 NF-κB诱导激酶(NIK)(一种MAPKKK)的激活可诱导IKKα二聚体复合物的磷酸化和激活,进而激活p100。这导致p100螯合释放p52 / RelB异二聚体。 p52 / RelB异二聚体易位至细胞核,以激活与许多细胞功能有关的靶基因,特别是细胞增殖,存活和先天免疫[202]。7.1.NF-κB途径的调节NF-κB途径已被证明在HCoV感染中起重要作用。重组SARS-CoV感染小鼠的肺中NF-κB被激活[203]。但是,在同一项研究中,随后用NF-κB抑制剂对这些感染的肺细胞进行处理不会影响病毒滴度,但会降低TNF,CCL2和CXCL2的表达,因此表明NF-κB对于SARS-CoV介导的SAR诱导是必不可少的。促炎细胞因子[203]。还显示出HCoV-229E介导外周血单核细胞(PBMC)中的IL8诱导,可被NF-κB抑制剂减弱[204]。 NF-κB的调节是通过HCoV的几种病毒蛋白介导的(图5)。7.1.1.结构蛋白在SARS-CoV中,已证明S,M,E和N结构蛋白会干扰NF-κB信号传导。最近的一项研究报道了用纯化和重组的SARS-CoV S蛋白处理过的PBMC中增强的核NF-κB活性。这些S蛋白处理过的细胞中IL8的合成和分泌可以被NF-κB抑制剂抑制,因此表明NF-κB调节了这些细胞中促炎性细胞因子的水平[204]。SARS-CoV S蛋白可能通过上游蛋白激酶C(PKC)同工酶PKCα的上调刺激NF-κB,因为S激活的ERK和JNK是PKC依赖性的[192]。尽管SARS-CoV E蛋白不能促进IL8的合成和分泌[204],但重组SARS-CoV中E蛋白的缺失会降低NF-κB的激活[203]。此外,SARS-CoV N蛋白的过表达在VeroE6细胞中以剂量依赖性方式显着增加了NF-κB荧光素酶的活性,但在Vero,HeLa和Huh-7细胞中却没有,这表明这种NF-κB的诱导可能是细胞-具体[205,206]。另一方面,免疫共沉淀实验表明,SARS-CoVM蛋白与IKKb物理结合,将其隔离在细胞质中,从而抑制了NF-κB的活化[207]。然而,尽管我们不能排除可能涉及某些其他结构或非结构性MERS-CoV蛋白的可能性,但MERS-CoV M蛋白并不影响由具有NF-κB结合位点的启动子控制的荧光素酶活性[133]。 。对于HCoV-OC43,除非受到TNFα的刺激,否则仅表达其N蛋白就无法激活NF-κB。 NF-κB活化的增强是通过HCoV-OC43 N与microRNA9的相互作用来抑制的NF-κB[208]。7.1.2.非结构蛋白和辅助蛋白以前,已证明SARS-CoV nsp1的过度表达可诱导NF-κB活化,而HCoV-229Ensp1则不能[138]。 NF-κB抑制剂的使用以剂量依赖的方式抑制SARS-CoV nsp1诱导的趋化因子表达[209]。在SARS-CoV和MERS-CoV的nsp3中发现的PLpro结构域可拮抗IFN和NF-κB的活性[139,210]。然而,在另一项研究中,SARS-CoV PLpro结构域并没有显着否定仙台病毒感染对NFκB依赖性基因表达的诱导[140]。然而,已经表明,SARS-CoV PLpro通过从IκBα去除K48连接的泛素化来抑制NF-κB的活化[211]。HCoV的其他辅助蛋白也已显示出干扰NF-κB信号传导的作用。 SARS-CoV 3a和7a蛋白的表达可显着诱导NF-κB依赖的荧光素酶活性。通过突变启动子上的NF-κB结合位点,可否定SARS-CoV 3a和7a蛋白增强IL8启动子的活性[192]。以前,在NFκB反应性启动子的控制下,表明MERS-CoV ORF4a的表达可抑制仙台病毒诱导的萤火虫荧光素酶活性,但不能抑制ORF4b和ORF5的表达[133]。然而,另一项研究表明,MERS-CoV ORF4b可适度减弱TNFα治疗诱导的NF-κB依赖的荧光素酶活性[212]。8.结论病毒与其宿主之间的关系是一件复杂的事情:来自病毒与宿主的众多因素都与病毒感染和相应的发病机理有关。在病毒感染期间,宿主必须通过采取多种防御机制来应对病毒。作为细胞内专性寄生虫,病毒还发展了各种劫持宿主机器的策略。在这篇综述中,我们首先展示了病毒因子如何操纵宿主细胞以加速其自身的复制周期和发病机理。我们还着重指出了多种细胞和病毒因素如何在长期的相互抗争中发挥作用。多年来,HCoV已被确定为影响人类的轻度呼吸道病原体。然而,正是SARS-CoV的出现使这些人类病毒成为研究领域的焦点。因此,当今大多数HCoV研究都与SARS-CoV有关。虽然最近的MERS-CoV暴发主要限于中东地区,但从过去两次暴发中的高死亡率可以看出,更多的新兴或重新出现的HCoV可能会威胁到全球公共卫生: SARS冠状病毒(10%)和MERS冠状病毒(35%)。因此,对所有HCoV的发病机理的研究将为抗病毒治疗剂和疫苗的开发获得更多见识。致谢:这项研究得到了新加坡国家研究基金会的竞争研究计划(CRP)资助(NRF-CRP8-2011-05)的部分支持,南阳的学术研究基金(AcRF)一级资助(RGT17 / 13)新加坡科技大学和教育部,以及新加坡教育部的AcRF Tier 2资助(ACR47 / 14)。作者贡献:Yvonne Xinyi Lim和Nyan Ling Ng撰写了这篇论文; James Tam。Tam和Liu Ding Xiang修改了手稿。参考文献(展示部分)1. 佩内(F.) A.Merlat; Vabret,A .;Rozenberg,F。 A. Buzyn; Dreyfus,F .;卡里乌(A.Freymuth,F .; Lebon,P.免疫受损患者的冠状病毒229E相关性肺炎。临床感染。 Dis。 2003,37,929–932。[CrossRef] [PubMed]2. 维杰根湖; E. Keyaerts; E.Moës; Maes,P .; Duson,G .; van Ranst,M.开发用于人冠状病毒OC43和229E绝对定量的一步式实时定量逆转录酶PCR分析方法。 J.临床。微生物。 2005,43,5452-5456。 [CrossRef] [PubMed]3. 琼斯,学士学位;格蕾丝·D;科克河;阿隆索,S。 Rushton,J .;说,M.Y .;麦基弗(D.穆图阿杨,J .;麦克德莫特,J。等。人畜共患病的出现与农业集约化和环境变化有关。进程Natl。学院科学美国,2013年,第21卷,第8399-8340页。 [CrossRef] [PubMed]4. Vander Hoek,L.人冠状病毒:它们引起什么?抗病毒那个2007,12,651–658。 [考研]5. Walsh2007,E.E .; Shin,J.H .;Falsey,A.R。人类冠状病毒229E和OC43感染在不同成年人群中的临床影响。 J.感染。 Dis。 2013,208,1634–1642。 [CrossRef] [PubMed]6. Gorse,G.J .; T.Z. O’Connor; Hall,S.L .; J.N.尼科尔(K.L.人冠状病毒和老年慢性阻塞性肺疾病的急性呼吸系统疾病。 J.感染。 Dis。 2009,199,847–857。 [CrossRef] [PubMed]7. N. Arbour;天,R。纽科姆,J。 Talbot,P.J.人类呼吸道冠状病毒的神经入侵。J.维罗尔2000,74,8913–8921。 [CrossRef][PubMed]8. N. Arbour;Ekandé,S.; G.Côté; Lachance,C .; Chagnon,F。塔迪厄,M。 N.R. Cashman;Talbot,P.J.人冠状病毒229E持续感染人少突胶质细胞和神经胶质细胞系。 J.维罗尔1999,73,3326-3337。 [考研]9. Jacomy,H.;弗拉戈索,G。阿尔玛赞(G.)华盛顿州Mushynski; Talbot,P.J.人冠状病毒OC43感染会诱发慢性脑炎,导致BALB / C小鼠残疾。病毒学,2006,349,335-346。[CrossRef] [PubMed]10. Vabret,A .; T. Mourez; Gouarin,S。 Petitjean,J.; Freymuth,F.法国诺曼底爆发冠状病毒OC43呼吸道感染。临床感染。 Dis。 2003,36,985–989。[CrossRef] [PubMed]11. Smuts,H.南非因急性呼吸道感染住院的婴儿的人类冠状病毒NL63感染。流感其他呼吸道。病毒2008,2,135-138。 [CrossRef] [PubMed]12. 格雷厄姆(R.L.); E.F.唐纳森; R.S.BaricSARS十年后:控制新兴冠状病毒的策略。纳特微生物牧师。 2013,11,836–848。 [CrossRef] [PubMed]13. 弗里曼,M。 Baric,R.严重急性呼吸系统综合症的发病机制和先天免疫调节机制。微生物。大声笑生物学修订版MMBR 2008,72,672-685。 [CrossRef] [PubMed]14. Peiris,J.S.M .;关Y袁凯严重的急性呼吸道综合症。 Nat.Med.2004,10,S88-S97。 [CrossRef][考研]15. 王敏;严敏;徐辉;梁伟;坎,B。郑宝陈华;郑慧;徐Y张娥等。 Palm Civet一家餐厅的SARS-CoV感染。新兴的。感染。 Dis。 2005,11,1860–1865。 [CrossRef] [PubMed]16. 胡乙; Ge,X .; WangL.-F.; Shi,Z。Bat是人类冠状病毒的起源。病毒。 J. 2015,12,221. [CrossRef] [PubMed]17. 金,Y。Cheon,S .;敏,C.-K .; Sohn,K.M .;康玉杰; Cha,Y.-J.;姜智智;韩,哈,纽约;金(G.等。在韩国爆发期间,与人类CD26亲和力降低的变异中东呼吸系统综合症冠状病毒的传播。 mBio 2016,7。[CrossRef] [PubMed]18. 奥本(I.K.) Tomczyk,S.M .; Al-Asmari,上午;班加尔(Banjar) Al-Mugti,H .;硕士Aloraini,M.S .; K.Z. Alkhaldi; E.L.; Almohammadi;阿拉木图(Brad) Gerber,S.I .;等。 2014年在吉达的中东呼吸综合征冠状病毒暴发—通往医疗机构的链接。N.Engl。 J. Med。2015,372,846–854。 [CrossRef][PubMed]19. 韩国传染病学会;大韩民国保健相关感染控制与预防协会。 2015年,大韩民国意外爆发的中东呼吸综合征冠状病毒感染。感染。化学药剂师。 2015,47,120–122。20. 硕士冠状病毒的分子生物学在病毒研究中的进展;学术出版社:马萨诸塞州,美国马萨诸塞州,2006年;第66卷,pp。 193–292。21. 麦克布赖德河;卑诗省菲尔丁严重急性呼吸系统综合症(SARS)-冠状病毒辅助蛋白在病毒发病中的作用。病毒2012,4,2902–2923。 [CrossRef] [PubMed]22. A.E. Gorbalenya;斯奈德,E.J .;斯潘,W.J.M.严重急性呼吸系统综合症冠状病毒系统发育:达成共识。 J.维罗尔2004,78,7863–7866。 [CrossRef] [PubMed]23. 科列斯尼科娃(L. W.Slenczka;布罗特,H。Klenk,H .; Becker,S.在SARS病例诊断中的电子显微镜检查。 Microsc。微肛门。 2003,9,438–439。24. G.Marsolais; Berthiaume,L .; E.DiFranco; Marois,P.通过电子显微镜快速诊断禽冠状病毒感染。能够。 J.比较中1971,35,285–288。 [考研]25. 刘德信;冯T.S .; Chong,K.K.-L .; Shukla,A .; Hilgenfeld,R. SARS-CoV和其他冠状病毒的辅助蛋白。抗病毒Res。 2014,109,97–109。 [CrossRef] [PubMed]26. Yeager,C.L .; R.A. Ashmun;英国威廉姆斯;卡德利基奥(C.B.) L.H. Shapiro;看着。;霍姆斯人氨基肽酶N是人冠状病毒229E的受体。自然1992,357,420-422。[CrossRef] [PubMed]27. 李伟;摩尔,新泽西州;瓦西里耶娃(N.隋J.黄锡基;马萨诸塞州伯恩; M. Somasundaran;沙利文(J.L.);卢祖里加(K.华盛顿州格林诺市;等。血管紧张素转化酶2是SARS冠状病毒的功能性受体。自然,2003,426,450-454。 [CrossRef] [PubMed]28. 李伟;隋J.黄,IC。库恩(J.H. Radoshitzky,S.R .;华盛顿州马拉斯科;崔H. Farzan,M.人冠状病毒NL63和严重急性呼吸系统综合症冠状病毒的S蛋白结合ACE2的重叠区域。病毒学2007,367,367–374。 [CrossRef] [PubMed]29. 吴克;李伟;彭庚; Li,F.与人类受体复合的NL63呼吸冠状病毒受体结合域的晶体结构。进程Natl。学院科学美国2009,106,19970–19974。 [CrossRef] [PubMed]30. 范多雷马伦(N. K.L. Miazgowicz; Milne-Price,S .; Bushmaker,T。罗伯逊,S。斯科特(D.)Kinne,J .;麦克莱伦(J.S.)朱建芒斯特通过其受体二肽基肽酶4限制中东呼吸综合征冠状病毒的宿主物种。J.Virol。 2014,88,9220–9232。 [CrossRef] [PubMed]31. 黄X. Dong,W .; A.Milewska; Golda,A .;齐Y;朱庆功;华盛顿州马拉斯科; R.S. Baric; A.C. Sims; Pyrc,K .;等。人类冠状病毒HKU1穗蛋白使用O-乙酰化唾液酸作为附着受体决定簇,并使用血凝素酯酶蛋白作为破坏受体的酶。 J.维罗尔2015,89,7202–7213。 [CrossRef] [PubMed]32. 北角巴特勒; Pewe,L .; K.Trandem;由人HCoV-OC43引起的Perlman,S.鼠脑炎具有广泛物种特异性的冠状病毒部分是免疫介导的。病毒学,2006,347,410–421。 [CrossRef] [PubMed]33. A.Zumla;陈建伟; E.I.Azhar;许,; Yuen,K.冠状病毒—药物发现和治疗选择。纳特Rev Disc Drug Discov。 2016,15,327–347。 [CrossRef] [PubMed]34. B.J.博世;华盛顿州的Bartelink。罗蒂尔(P.J.M.)组织蛋白酶L在功能上切割严重急性呼吸系统综合症冠状病毒I类融合蛋白,而不是毗邻融合肽。 J.维罗尔2008,82,8887-8890。 [CrossRef] [PubMed]35. 钱正; S.R. Dominguez;霍姆斯人中东呼吸综合征冠状病毒(MERS-CoV)的穗状糖蛋白在病毒进入和合胞体形成中的作用。公共科学学报2013,8,e76469。 [CrossRef] [PubMed]36. 西蒙斯,G .; D.N. Gosalia;伦纳坎普(Rennekamp)里夫斯,法学博士; Diamond,S.L .; Bates,P.组织蛋白酶L的抑制剂可防止严重的急性呼吸系统综合症冠状病毒进入。进程Natl。学院科学美国,2005年,102,11876-11881。 [CrossRef] [PubMed]37. Bertram,S。 Dijkman,R .;哈布赞(M. Heurich,A .; Gierer,S。 Glowacka,我。威尔士(K.) M. Winkler;施耐德,H。Hofmann-Winkler,H .;等。 TMPRSS2激活人类冠状病毒229E,使组织蛋白酶不依赖宿主细胞进入,并在呼吸道上皮的病毒靶细胞中表达。 J.维罗尔2013,87,6150-6160。 [CrossRef] [PubMed]38. Bertram,S .; Glowacka,我。 Müller,硕士。薰衣草,H。 Gnirss,K .;尼尔米尔,我。 Niemeyer,D .;他,Y。西蒙斯,G .; Drosten,C .;等。人气道胰蛋白酶样蛋白酶对严重急性呼吸系统综合症冠状病毒刺突蛋白的裂解和激活。 J.维罗尔2011,85,13363–13372。 [CrossRef] [PubMed]39. J.K. Millet;惠特克(G.R.)经过弗林蛋白酶介导的穗蛋白激活两步后,中东呼吸综合征冠状病毒进入宿主细胞。进程Natl。学院科学美国,2014年,111,15214–15219。 [CrossRef] [PubMed]40. 黄,IC。哥伦比亚贝利;Weyer,J.L .; Radoshitzky,S.R .;贝克尔(M.M.);蒋俊杰;黄铜,A.L .;艾哈迈德(Ahmed)池X.董琳;等。 IFITM介导的丝状病毒,SARS冠状病毒和甲型流感病毒限制的独特模式。 PLoS Pathog。 2011,7,e1001258。 [CrossRef] [PubMed]41. 李K马科斯扬(RM。郑玉男; O.戈尔菲托; Bungart,B。李敏丁,;他,Y。梁昌Lee J.C .;等。 IFITM蛋白可限制病毒膜半融合。 PLoS Pathog。 2013,9,e1003124。 [CrossRef] [PubMed]42. 罗,陈青;陈建陈K沉X. Jiang,H。SARS冠状病毒的核衣壳蛋白与人细胞异种核糖核糖核蛋白A1具有很高的结合亲和力。 FEBS Lett。 2005,579,2623–2628。 [CrossRef] [PubMed]43. 南达(S.K.) Leibowitz,J.L.线粒体乌头酸酶与小鼠的31个非翻译区结合肝炎病毒基因组。 J.维罗尔2001,75,3352–3362。 [CrossRef] [PubMed]44. Wu,C.-H .;陈平杰;是的S.-H.核仁磷酸化和RNA解旋酶DDX1的招聘使冠状病毒从不连续转录过渡到连续转录。细胞宿主微生物。 2014,16,462–472。 [CrossRef] [PubMed]45. Tan,Y.W .; Hong,W.;刘德祥冠状病毒RNA 51非翻译区与锌指的结合CCHC型和RNA结合基序1增强病毒复制和转录。核酸研究。 2012年40,5065–5077。 [CrossRef][PubMed]46. 纽曼(B.W.)吻,G。 A.H.昆定; Bhella,D .; Baksh,M.F .; Connelly,S.; Droese,B .; J.P.克劳斯; Makino,S .; Sawicki,S.G .;等。冠状病毒装配和形态中M蛋白的结构分析。J.结构。生物学2011,174,11-22。[CrossRef] [PubMed]47. 罗,吴丹;沉丙;陈K沉X.严重急性呼吸系统综合症冠状病毒膜蛋白主要通过静电吸引作用通过羧基末端与核衣壳蛋白相互作用。诠释J.生物化学。细胞生物学。 2006,38,589–599。 [CrossRef] [PubMed]48. Wong H.H .;库玛尔; Tay,F.P.L .; D.Moreau;刘德信; Bard,F.全基因组筛选揭示了冠状病毒从内体退出的含缬氨酸的蛋白质需求。 J.维罗尔2015,89,11116-11128。 [CrossRef] [PubMed]49. J.F.R. A.H. Wyllie;柯里(A.R.)凋亡:一种基本的生物现象,对组织动力学具有广泛的影响。 Br。 J. Cancer 1972,26,239–257。 [CrossRef] [PubMed]50. Boulares,A.H .;A.G. Yakovlev;伊万诺娃(Ivanova)B.A. Stoica;王庚; Iyer,S .; Smulson,M.聚(ADP-核糖)聚合酶(PARP)切割在细胞凋亡中的作用:抗Caspase 3的parp突变体增加了转染细胞的凋亡率。 J.Biol。化学1999,274,22932-22940。 [CrossRef] [PubMed]51. 濑川市;仓田Yanagihashi,Y .; T.R. Brummelkamp;F.松田; Nagata,S.半胱天冬酶介导的磷脂翻转酶裂解,引起凋亡的磷脂酰丝氨酸暴露。 Science 2014,344,1164–1168。 [CrossRef] [PubMed]52. Walczak,H.;克拉默CD95(APO-1 /Fas)和TRAIL(APO-2L)细胞凋亡系统。经验细胞水库。 2000,256,58–66。[CrossRef][PubMed]53. 班德(L.M.)摩根(Morgan)托马斯(Thomas L.R.)刘志国; Thorburn,A.衔接蛋白TRADD激活细胞核和细胞质凋亡的独特机制。细胞死亡差异。 2005,12,473–481。 [CrossRef] [PubMed]54. Stennicke,H.R .; Jürgensmeier,J.M .; Shin,H。 Q.Deveraux;沃尔夫,B.B .;杨X.周青;Ellerby,H.M .; Ellerby,L.M .;布雷德森等。前胱天蛋白酶-3是胱天蛋白酶8的主要生理指标。 J.Biol。化学1998,273,27084–27090。 [CrossRef] [PubMed]点击:查看更多医学文章 试用免费翻译功能 人类冠状病毒:病毒与宿主相互作用的综述(中) 免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:mdpi
2021-01-22 17:00:50

人类冠状病毒:病毒与宿主相互作用的综述(上)与TLR不同,RLR和NLR普遍存在。 RLR是一个胞质受体家族,由三个成员组成:视黄酸诱导基因I(RIG-1),黑素瘤分化相关因子5(MDA5)和遗传与生理学实验室2(LGP2)。 RIG-1和MDA5在其N端具有两个胱天蛋白酶招募域(CARD),一个DExD / H-box RNA解旋酶域(其中x可以是任何氨基酸)和一个在C端的阻遏域(RD)。另一方面,LGP2缺少CARD域[112],并且它可以调节RIG-I和MDA5。积极或消极[113–115]。 RIG-I识别病毒基因组中存在的51-三磷酸部分RNA,以及通过病毒基因组互补末端的自退火形成的双链“panhandle”结构[116,117]。相反,MDA5通常会检测到更长的dsRNA序列。 RIG-1和MDA5与病毒RNA的结合会导致构象变化,从而暴露CARD域。募集了一种定位于线粒体和过氧化物酶体的衔接子蛋白,称为线粒体抗病毒信号转导适配器,MAVS。然后,MAVS激活转录因子,例如干扰素调节因子1(IRF1),IRF3和NF-κB,以触发干扰素(IFN)和促炎性细胞因子
2021-01-21 17:30:55

人类冠状病毒:病毒与宿主相互作用的综述Yvonne X in一l IM, y按L ing ng, James P. ta们and ding X Ian GL IU *南洋理工大学生物科学学院,南洋大道60号,新加坡637551;新加坡; YVON0016@e.ntu.edu.sg(Y.X.L.); S150004@e.ntu.edu.sg(Y.L.N.); JPTAM@ntu.edu.sg(J.P.T.)收到:2016年6月8日;接受:2016年7月18日;发布时间:2016年7月25日摘要:人类冠状病毒(HCoV)是已知的与一系列呼吸结果相关的呼吸道病原体。在过去的14年中,重度急性呼吸综合征冠状病毒(SARS-CoV)和中东呼吸综合征冠状病毒(MERS-CoV)的发作使HCoV在人类中具有很高的致病性,已成为研究界的关注焦点。HCoV-宿主相互作用的研究为我们对HCoV发病机理的理解做出了广泛贡献。在这篇综述中,我们讨论了宿主细胞因子的最新发现,这些因子可能被HCoV利用以促进其自身的复制周期。我们还讨论了各种细胞过程,例如细胞凋亡,先天免疫,内质网应激反应,有丝分裂原激活的蛋白激酶(MAPK)途径和可能由HCoV调节的核因子κB(NF-κB)途径。 关键词:人冠状病毒;病毒与宿主的相互作用;细胞凋亡先天免疫;内质网应激MAPK;核因子κB 1. 介绍人冠状病毒(HCoV)代表与严重性不同的多种呼吸系统疾病相关的主要冠状病毒(CoV),包括普通感冒,肺炎和支气管炎[1]。如今,由于HCoV具有高的基因组核苷酸取代率和重组能力,因此被公认为发展最快的病毒之一[2]。近年来,HCoV的进化也受到诸如城市化和家禽养殖等因素的推动。这些已允许物种的频繁混合,并促进了这些病毒的物种屏障和基因组重组的穿越[3]。迄今为止,已鉴定出六种已知的HCoV,即HCoV-229E,HCoV-NL63,HCoV-OC43,HCoV-HKU1,严重急性呼吸综合征冠状病毒(SARS-CoV)和中东呼吸综合征冠状病毒(MERS-CoV);其中,四种HCoV(HCoV-229E,HCoV-NL63,HCoV-OC43和HCoV-HKU1)在全球范围内流通,约占人类普通感冒感染的三分之一[4]。在严重的情况下,这四种HCoV可以引起危及生命的肺炎和细支气管炎,特别是在老年人,儿童和免疫功能低下的患者中[1,5,6]。除呼吸系统疾病外,它们还可能引起肠道和神经系统疾病[7-11]。SARS-CoV最初于2002-2003年在中国广东出现,是一种典型的非典型肺炎,其特征是发烧,头痛以及随后出现的咳嗽和肺炎等呼吸道症状,后来可能发展为危及生命的呼吸衰竭和急性呼吸窘迫综合征[ 12]。它在人类中具有很高的传播性,并迅速传播到29个国家/地区,感染了8000多人,死亡率约为10%[13,14]。最初,棕榈果被认为是病毒的天然贮藏库[15]。但是,随后的系统发育研究指出,SARS-CoV的蝙蝠起源是基于蝙蝠中发现的SARS样病毒序列[16]。 2012年,沙特阿拉伯出现了MERS-CoV流行病,其临床症状与SARS-CoV类似,但是死亡率要高得多,约为35%[17]。与表现出超级传播事件的SARS-CoV不同,MERS-CoV的传播受地域限制[12]。实际上,报告的MERS-CoV病例通常源于中东国家内的暴发或近期到该地区的旅行[18,19]。分类学,基因组结构和形态CoV是冠状病毒科下的一组大型包膜RNA病毒。冠状病毒科与Artierivirdae和Roniviridae一起归类于Nidovirale顺序[20]。根据国际病毒分类学委员会的建议,基于整个病毒基因组的序列比较,冠状病毒进一步分为四个主要属,α-,β-,γ-和三角冠状病毒[21,22]。这些冠状病毒可感染多种宿主,包括禽,猪和人类。已鉴定出HCoV属于Alpha或Beta冠状病毒属,包括Alphacoronaviruses,HCoV-229E和HCoV-NL63以及Betacoronaviruses,HCoV-HKU1,SARS-CoV,MERS-CoV和HCoV-OC43(表1)。在电子显微镜下,冠状病毒病毒颗粒看起来大致呈球形或中等多形性,由刺突蛋白(S)形成独特的“棍状”突起[23,24]。在病毒体内部有一个螺旋对称的核衣壳,它包裹着一个巨大的单链和正向RNA病毒基因组,其大小约为26至32千个碱基[20]。正性病毒基因组RNA充当信使RNA(mRNA),包括51个末端帽结构和31个poly A尾巴。这种基因组RNA在病毒生命周期中具有三种功能:(1)作为感染周期的初始RNA; (2)作为复制和转录的模板; (3)作为包装入子代病毒的底物。复制酶-转录酶是唯一从基因组翻译的蛋白质,而所有下游开放阅读框的病毒产物均来自亚基因组mRNA。在所有冠状病毒中,复制酶基因约占基因组的51分之三,由两个重叠的开放阅读框(ORF),ORF1a和ORF1b组成,它们编码16个非结构蛋白。 CoV基因组RNA的最后三分之一编码四个结构蛋白基因的CoV规范集,顺序为尖峰(S),包膜(E),膜(M)和核衣壳(N)。此外,还沿着结构蛋白基因散布了一些辅助ORF。CoV物种的数量和位置各不相同[25](图1)。图1.人类冠状病毒(HCoV)的基因组组织。 HCoV基因组的大小范围约为26至32千个碱基(kb),如刻度上方的黑线所示。冠状病毒(CoV)基因组通常按51-ORF1a-ORF1b-S-E-M-N-31的顺序排列。重叠的开放阅读框(ORF)ORF1a和ORF1b占冠状病毒基因组的三分之二,该基因组编码了病毒RNA合成所需的所有病毒成分。基因组31末端的另外三分之一编码一组结构蛋白(橙色)和非结构蛋白(绿色)。表1.人冠状病毒的分类。冠状病毒科 菌株发现细胞受体主办参考文献冠状病毒HCoV-229E 1966人氨基肽酶N(CD13)蝙蝠[1,2,21]肝炎病毒-NL632004ACE2棕榈果,蝙蝠[3,21]冠状病毒OC4319679-O-乙酰化唾液酸牛[4,5]β冠状病毒HcoV-HKU1 20059-O-乙酰化唾液酸老鼠[6,7]SARS冠状病毒2003ACE2棕榈果,蝙蝠[8,19,21]冠状病毒2012DPP4蝙蝠,骆驼[9] 2. 宿主因素参与病毒复制和发病机理作为细胞内专性寄生虫,HCoV利用宿主细胞机制进行自身复制和传播。由于病毒与宿主之间的相互作用是疾病的基础,因此有关其相互作用的知识引起了极大的研究兴趣。在这里,我们描述了细胞在冠状病毒感染周期中的作用的已知情况:附着;进入宿主细胞;复制酶-转录酶的翻译;基因组复制和mRNA转录;新包装的病毒体的组装和萌芽(图2)。图2.冠状病毒复制周期。冠状病毒感染始于刺突蛋白(S)的S1结构域与其同源受体的附着。这驱动了S中S2亚基的构象变化,促进了病毒和细胞质膜的融合。核衣壳释放到细胞质后,病毒gRNA通过核糖体移码翻译产生多蛋白pp1a和pp1ab。 pp1a和pp1ab由宿主和病毒蛋白酶进行蛋白水解处理,生成16个非结构蛋白(NSP),然后将其组装形成复制酶-聚合酶。复制酶-聚合酶参与冠状病毒的复制,在该过程中基因组RNA被复制,亚基因组RNA被转录并翻译以形成结构蛋白。产生的病毒产物将在ERGIC中组装,并以光滑壁囊泡的形式发芽到质膜,通过胞吐作用排出。促进感染和抑制感染的宿主因子分别以绿色和红色突出显示。 APN,氨肽酶N; ACE2,血管紧张素转化酶2; DPP4,二肽基肽酶4; 9-O-Ac唾液酸,9-O-乙酰化唾液酸; IFITM,干扰素诱导的跨膜蛋白; ATP1A1,ATPase,Na + / K +转运,α1多肽; HnRNP A1,异核核糖核蛋白A1; MADP1,锌指CCHC型和RNA结合基序1; DDX1,ATP依赖性RNA解旋酶; PCBP1 / 2,Poly r(C)结合蛋白1/2;PABP,Poly A结合蛋白;COPB2,Coatomer蛋白复合物,β2亚基(β素); GAPDH,甘油醛3-磷酸脱氢酶; ERGIC,内质网高尔基体中间隔间;ER,内质网;VCP,含缬氨酸的蛋白。 2.1. 冠状病毒附着和进入CoV感染是通过刺突(S)蛋白附着于特定宿主细胞受体而引发的。宿主受体是病毒的致病性,组织嗜性和宿主范围的主要决定因素。 S蛋白包含两个结构域:S1和S2。 S1结构域及其同源受体之间的相互作用触发S蛋白的构象变化,然后通过S2结构域促进病毒和细胞膜之间的膜融合。今天,所有HCoV使用的主要宿主细胞受体是已知的:HCoV-229E产生的氨肽酶N[26],SARS-CoV产生的血管紧张素转换酶2(ACE2)[27]和HCoV-NL63[28,29],二肽MERS-CoV的肽酶4(DPP4)[30]和HCoV-OC43和HCoV-HKU1的9-O-乙酰化唾液酸[31,32]。除了常规的内体进入途径外,某些冠状病毒也可能进入通过非内体途径,或两者结合的细胞。细胞环境中的低pH值和内体半胱氨酸蛋白酶组织蛋白酶可能有助于促进膜融合和内体CoV细胞进入[33]。最近的证据支持组织蛋白酶L在SARS-CoV和MERS-CoV进入中的作用[34-36]。其他宿主蛋白酶,例如跨膜蛋白酶丝氨酸2(TMPRSS2)和气道胰蛋白酶样蛋白酶TMPRSS11D,也可以在HCoV-229E和SARS期间进行S1 /S2裂解以激活S蛋白,使非蛋白体病毒进入细胞质膜。 -CoV感染[37,38]。此外,MERS-CoV也被弗林蛋白酶激活,弗林蛋白酶是一种丝氨酸内肽酶,已与其他RNA病毒的细胞进入和病毒流出过程中的S1/ S2裂解有关[39]。许多宿主细胞还利用其自身的因素来限制病毒的进入。利用细胞培养系统和假型病毒,许多研究小组确定了干扰素诱导型跨膜蛋白(IFITM)家族,它们可以抑制全局循环的HCoV-229E和HCoV-NL63 S蛋白介导的进入,以及高致病性的SARS-CoV和MERS -CoV [12,40]。尽管IFITM的作用方式仍然难以捉摸,但一些研究小组进行的细胞间融合试验表明,IFITM3通过调节宿主膜的流动性来防止病毒包膜与质膜或内体膜融合,从而阻止了包膜病毒的进入[ 41]。2.2. 冠状病毒复制在病毒核衣壳释放并脱去细胞质后,CoV复制开始于将ORF 1a和1b翻译成多蛋白pp1a(4382个氨基酸)和pp1ab(7073个氨基酸)。在这里,下游的ORF1b通过核糖体移码机制进行翻译,在这种机制中,翻译的核糖体沿´1方向从ORF1a阅读框转移到ORF1b阅读框。通过两个RNA元件(51-UUUAAAC-31七核苷酸滑序列和RNA假结结构)可以实现重新定位。随后,将多蛋白pp1a和pp1ab切割成至少15 nsp,它们组装并形成复制转录复合体。通过复制酶聚合酶的组装,转录基因组RNA的全长正链,形成全长负链模板,用于合成新的基因组RNA和重叠的亚基因组负链模板。然后将这些亚基因组mRNA转录并翻译以产生结构蛋白和辅助蛋白。已经发现几个异源核糖核蛋白(hnRNA)家族成员(hnRNPA1,PTB,SYN-CRYP)对于有效的RNA复制是必不可少的[42]。还建议其他RNA结合蛋白在冠状病毒复制中发挥作用,例如间-α固醇酶和聚A结合蛋白(PABP),DDX1,PCBP1 / 2和锌指CCHC型和RNA结合基序1 (MADP1)[43-45]。2.3. 冠状病毒组装和出口随着新基因组RNA和结构组分的积累,病毒体的组装很快得以实现。在感染周期的这个阶段,含有基因组RNA的螺旋核衣壳与其他病毒结构蛋白(S,E和M蛋白)相互作用形成组装的病毒体。 CoV颗粒的组装是通过从内质网到高尔基体的分泌途径中早期的膜状螺旋核衣壳出芽而完成的。中间隔层(ERGIC)。很少探讨宿主在感染周期此阶段的贡献。目前,已知M蛋白通过在组装位点选择和组织病毒包膜成分并介导与核衣壳的相互作用来使病毒粒子出芽来协调整个组装过程[46]。M蛋白与不同的病毒结构蛋白(例如E蛋白)相互作用,组装成成熟病毒。这种相互作用产生了病毒粒子包膜的支架,并诱导了M蛋白修饰的膜以及与S蛋白的萌芽和释放,从而将刺突组装到病毒包膜中[46,47]。组装和出芽后,病毒粒子在囊泡中运输,并最终通过胞吐作用释放。在最近的一项研究中,抑制含缬氨酸的蛋白质(VCP / p97)导致病毒在感染性支气管炎病毒(IBV)的早期内体中蓄积,表明VCP在负载病毒的内体成熟中发挥了作用[48]。3. 人冠状病毒感染和凋亡凋亡是程序性细胞死亡的过程,其受到严格调节和消炎。当细胞发生凋亡时,它们表现出特定的标志,例如细胞萎缩,广泛的质膜起泡,核变性,DNA断裂和质膜的不对称分布[49-51]。迄今为止,已经建立了两种主要的细胞凋亡机制-外在途径和内在途径。外在途径是通过细胞外死亡配体(例如Fas配体(FasL)和TNF受体相关的凋亡诱导配体(TRAIL))与来自肿瘤坏死因子(TNF)超家族的死亡受体的结合而启动的[52] ]。然后,这些死亡受体募集各种死亡衔接蛋白,例如Fas相关的死亡结构域蛋白(FADD)[53],以及启动子蛋白酶8和10形成诱导死亡的信号复合物(DISC)[54,55]。因此,两个启动子原蛋白酶被切割成其活性形式并诱导信号传导级联,最终激活效应子胱天蛋白酶3和7。另一方面,内在途径在细胞内部发生,并涉及线粒体外膜通透性(MOMP)的变化。 )基于促凋亡和抗凋亡B细胞淋巴瘤2(Bcl2)家族蛋白质的比率(图3)。增强的MOMP会导致促凋亡因子(例如细胞色素c)的释放,从而激活启动子胱天蛋白酶9。像外在途径一样,内在途径中的启动子胱天蛋白酶9的激活会导致蛋白水解裂解效应子胱天蛋白酶3和7,进而分解许多蛋白酶。凋亡必不可少的关键细胞蛋白[56]。当Bid(一种促凋亡的Bcl2家族蛋白)直接被caspase 8裂解时,甚至在效应子caspase激活之前,两条途径之间的融合也可能发生。在病毒感染期间,凋亡被诱导为宿主抗病毒反应之一,以限制病毒复制和生产。许多病毒已经进化出不同的策略来破坏细胞凋亡[58]。例如,某些病毒编码充当Bcl2家族蛋白同源物的病毒蛋白[59]。另外,病毒可能会通过其他分子途径,例如有丝分裂原活化蛋白(MAPK)和核因子κB(NF-κB)途径,直接或间接地建立调节Bcl2家族蛋白或半胱天冬酶激活的机制[60-65]。有趣的是,某些病毒可能会参与凋亡机制以进行有效的病毒感染。例如,甲病毒和黄病毒含有富含磷脂酰丝氨酸的病毒膜,以模仿凋亡细胞以促进病毒进入[66]。3.1. 细胞趋向与凋亡由于HCoV是已知可感染源自呼吸道的组织培养物和细胞系的呼吸道病原体,因此这些病毒也可能感染其他组织培养物和细胞系。这些组织和细胞的感染可能会诱导细胞凋亡[67,68]。然而,尽管HCoV在感染期间主要靶向呼吸道,但它们还与多种细胞类型的凋亡诱导相关,包括肠道粘膜细胞,肾小管细胞和神经元细胞[69-74]。对SARS-CoV感染组织的尸检研究表明,在肺,脾和甲状腺中诱导了细胞凋亡[75]。还显示出HCoV会感染免疫系统并诱导免疫细胞(例如巨噬细胞,单核细胞,T淋巴细胞和树突状细胞)凋亡[69,76-79]。因为这些免疫细胞与先天性和获得性免疫的激活有关,有理由推测,大量消除这些细胞可能是抑制宿主先天性和适应性免疫反应的一种病毒策略。在最近的研究中,据报道HCoV-229E感染导致大量CPE和树突状细胞死亡[80]。由于树突状细胞遍布我们的整个身体,因此有可能将它们用作促进病毒传播和损害我们的免疫系统的媒介[80,81]。 图3. Bcl2蛋白家族对MOMP的调节。 (a)Bcl2蛋白质家族根据其功能和Bcl2同源性(BH)结构域的数量分为三大类。生存前的Bcl2样家族成员(Bcl2,B细胞超大淋巴瘤(Bcl-XL),骨髓细胞白血病(Mcl1))包含所有四个BH结构域,并且具有抗凋亡作用。第二类称为Bcl2相关X(BAX)样蛋白,包括BAX和Bcl2同源拮抗剂杀手(BAK),具有促凋亡作用,并且缺少BH4结构域。最后,第三类称为仅BH3蛋白(Bid,与Bcl2相关的死亡启动子(Bad)和p53上调的细胞凋亡调节剂(PUMA)),仅包含BH3结构域,并且具有促凋亡作用。 (b)提出了两种模型来解释Bcl2家族蛋白在MOMP中的作用-间接激活剂模型和直接激活剂-抑制剂模型[11]。在间接激活剂模型中,抗凋亡的Bcl2样蛋白抑制Bax-Bak孔复合物插入线粒体,从而促进MOMP和细胞色素c的释放。但是,当仅BH3的蛋白被激活超过某个阈值时,Bcl2样蛋白的抑制作用就会被破坏。在直接激活物-抑制剂模型中,仅BH3蛋白充当直接激活物,以诱导Bak-Bak插入线粒体外膜。这些仅BH3的蛋白质可以被类Bcl2的蛋白质抑制,而后者又可以被另一类仅BH3的蛋白质抑制。该图是对[12]的修改。3.2. 细胞凋亡的分子机制在分子水平上,据报道,HCoV感染可通过多种机制触发细胞凋亡。SARS-CoV诱导的凋亡显示为caspase依赖性的,并可能被caspase抑制剂Z-VAD-FMK或Bcl2的过表达抑制[82,83]。尽管病毒复制是诱导细胞凋亡所必需的[83],但细胞凋亡并不影响SARS-CoV的病毒复制动力学[82]。另一方面,MERS-CoV感染原代T淋巴细胞可诱导DNA片段化并激活caspase8和9激活,表明外部和内部途径均被激活。与SARS-CoV感染不同,MERS-CoV复制对于诱导受感染的T淋巴细胞凋亡不是必需的[79]。病原性较低的细胞也可以诱导细胞凋亡芯片数据证实,HCoVs菌株在HCoV-229E感染期间Bcl2家族成员的促凋亡和抗凋亡基因表达发生了显着变化[84]。 HCoV-OC43的感染可促进人神经元细胞中BAX易位至线粒体[74]。尽管胱天蛋白酶3和9在感染HCoV-OC43的鼠和人神经元细胞中被激活[9,74],但添加泛半胱天冬酶抑制剂Z-VAD-FMK和半胱天冬酶9抑制剂Z-LEHD-FMK不会影响这些感染的神经元细胞的活力,表明由HCoV-OC43诱导的程序性细胞死亡可能与胱天蛋白酶无关[74]。这突出了在HCoV感染中诱导非经典程序性细胞死亡机制的可能性。尽管在SARS-CoV中仅对此进行了研究,但HCoV感染过程中的凋亡机制很可能受病毒蛋白操纵(图4)。特别是,SARS-CoV S,N,E,M,ORF-6、7a和9b蛋白已显示在其宿主细胞中具有促凋亡功能[77,85-91]。 SARS-CoV E蛋白和7a蛋白的表达通过将抗凋亡的Bcl-XL蛋白螯合到内质网(ER)膜而促进了线粒体介导的细胞凋亡[77,92]。 SARS-CoV M蛋白也高度促凋亡,并介导半胱天冬酶8和9的激活[90]。此外,已显示HCoV-OC43野生型S蛋白在人神经元NT2-N和LA-N-5细胞系中诱导未折叠的蛋白应答(UPR),这可能导致细胞凋亡[93]。重组HCoV-OC43在其S蛋白处具有点突变,比野生型病毒诱导的caspase 3活化和核碎裂作用更强[93]。有趣的是,SARS-CoV N和9b蛋白的定位与caspase依赖性细胞凋亡的诱导有关[89,94]。这一发现为病毒蛋白的亚细胞定位和胱天蛋白酶激活之间的联系开辟了新的视角,而胱天蛋白酶激活是由HCoV调节细胞凋亡的一种方式。图4. HCoVs激活细胞凋亡。死亡配体与死亡受体的结合诱导胱天蛋白酶8激活,继而激活效应子胱天蛋白酶3和7以刺激细胞凋亡。另一方面,内在途径受促凋亡和抗凋亡的Bcl2家族蛋白(如Bcl-XL,Bcl2,Bax和Bak)调控,以诱导MOMP。随后由增强型MOMP引起的caspase 9激活刺激了caspase 3和7激活。在HCoV感染过程中,病毒或特定病毒蛋白(橙黄色方框)靶向外部和固有凋亡信号通路的多个阶段。4. 人冠状病毒感染和先天免疫当细胞暴露于病原体(如病毒)时,免疫应答以宿主防御的形式被诱导。在病原体暴露期间,以细胞类型依赖性方式调节免疫应答。在产生适应性免疫系统之前,先天免疫是抵御病毒的第一道防线。宿主和病毒都可以操纵先天免疫机制作为防御或逃避策略的一种形式[95,96]。 4.1. 模式识别受体免疫系统中的细胞通过几种识别策略来检测病毒病原体。其中,特征最清楚的是模式识别受体(PRR),其通过称为病原体相关分子模式(PAMPs)的进化保守结构与各种微生物病原体结合。 PRR主要分为三类,即Toll样受体(TLR),视黄酸诱导型基因I(RIG-1)样受体(RLR)和核苷酸寡聚化域(NOD)样受体(NLR)。TLR是一种I型跨膜蛋白,位于细胞表面或内体囊泡中。它们的富含亮氨酸的重复序列(LRR)域介导了对PAMP的识别以及来自细菌,真菌和病毒等各种来源的损伤相关分子模式(DAMP)[97]。 TLR的激活主要发生在抗原呈递细胞中,例如树突状细胞(DC),巨噬细胞,单核细胞和B细胞。在人类的10种已知TLR中,发现TLR2、3、4、7和9与病毒检测有关[98,99]。 TLR3识别双链RNA(dsRNA),这是病毒RNA复制过程中产生的复制中间体[100]。TLR7和8检测单链RNA(ssRNA),TLR9识别DNA病毒中存在的未甲基化CpG DNA [101-103]。除核酸外,其他TLR(例如TLR2和4)还可以检测到病毒蛋白,例如呼吸道合胞病毒(RSV),肝炎病毒,麻疹病毒和人类免疫缺陷病毒[104-107]。识别病毒成分后,TLR募集含有Toll /白介素1受体(TIR)的信号转接头分子,例如MyD88(髓样分化主要反应蛋白88)和含有TIR域的衔接子诱导干扰素-β(TRIF)[108–110]。然后,MyD88和TRIF刺激MAPK和NF-κB途径,以增强IFN和促炎性细胞因子的产生[111]。点击:病毒与宿主相互作用的综述(中) 查看更多医学文章 使用双语翻译功能免责声明:福昕翻译只充当翻译功能,此文内容及相关信息仅为传递更多信息之目的,仅代表作者个人观点,与本网站无关,版权归原始网站所有。仅供读者参考,并请自行核实相关内容。若需要浏览原文、下载参考文献等,请自行搜索文中提到的原文网站进行阅读。来源于:mdpi
2021-01-21 17:30:35
